Digestión, absorción y metabolismo de los lípidos en monogástricos y rumiantes
Publicado: 13 de mayo de 2020
Por: Aníbal Fernández Mayer. Técnico del INTA EEA Bordenave. Centro Regional Buenos Aires Sur (CERBAS). M Sc., Doctor y Post-Doc en Ciencias Veterinarias especialista en Nutrición Animal (Univ. Agraria La Habana, CUBA)
Son sustancias orgánicas insolubles en agua (hidrófobas) y solubles en solventes orgánicos como éter, benceno y cloroformo. En el análisis inmediato de los alimentos se incluyen en la fracción denominada Extracto etéreo.
Clasificación.
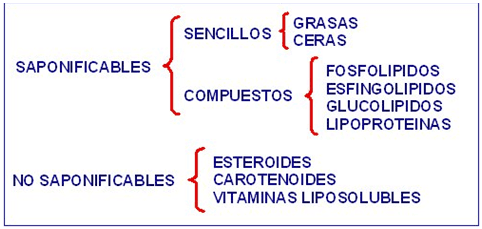
1. Lípidos simples saponificables.
A. Grasas.
Se trata de sustancias incoloras, inodoras e insípidas excepto cuando se enrancian que adquieren tonalidades más oscuras y amarillentas y modifican su olor y sabor. Las grasas o triglicéridos son esteres de la glicerina (o glicerol) con ácidos grasos.
En presencia de un álcali sufren el proceso de saponificación que consiste en la hidrólisis de la grasa formándose glicerol y liberándose los ácidos grasos que se unen al álcali formando una sal alcalina hidrosoluble (jabón). En el organismo la saponificación se produce merced a las lipasas segregadas por el páncreas.
Las grasas son sustancias hidrófobas (no solubles en agua) pero pueden formar emulsiones como, por ejemplo, las que se encuentran en la leche.
La mayor parte de las grasas alimentarias se suministran en forma de triacilglicéridos, que se deben hidrolizar para dar ácidos grasos (AG) y monoacilglicéridos (o glicerol) antes de ser absorbidos.
La digestión de las grasas se produce de forma eficaz y casi completa en el intestino delgado por secreción pancreática de lipasas. El estómago interviene en el proceso de digestión de las grasas debido a su acción agitadora, que ayuda a crear emulsiones.
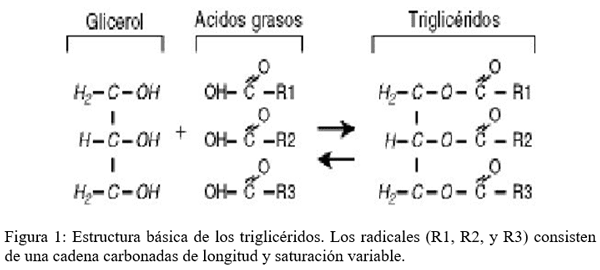
Las grasas que entran en el intestino se mezclan con la bilis y posteriormente se emulsionan.
La lipasa pancreática cataliza la hidrólisis de los ácidos grasos de las posiciones 1 y 3, generando 2-monoacilglicéridos.
La lipasa de lipoproteínas, que se encuentra en la pared interior de los capilares sanguíneos, hidroliza los triglicéridos, liberando AG. Estos AG entran en el tejido adiposo, donde se almacenan, y en los músculos, donde se utilizan como combustible.
Los ésteres del colesterol son hidrolizados por la hidrolasa de ésteres de colesterol pancreática.
Los ácidos grasos libres y los monoglicéridos (o glicerol) son absorbidos por los enterocitos, las célulasde la pared intestinal.
Los AGinferiores a 14 átomos de carbono entran directamente en el sistema de la vena porta y son transportados hacia el hígado.
Los AG con14 o más carbonos se vuelven a esterificar dentro del enterocito y entran en circulación a través de la ruta linfática en forma de quilomicrones. Sin embargo, la ruta de la vena porta también es una ruta de absorción de los AG.
Los quilomicrones son partículas lipoproteicas que proceden de las grasas alimentarias y son empaquetadas por las células de la mucosa. Entran en el torrente sanguíneo a través de los vasos linfáticos.
La mayoría de los ácidos poliinsaturados se comportan como ácidos grasos esenciales.
Las grasas vegetales, de los peces y las aviares tienen mayor grado de insaturación que las de los mamíferos. También dentro de un mismo animal la composición de la grasa varía, por ejemplo, la grasa corporal de una vaca está más saturada (más consistente), que la de la leche.
La grasa de la leche está integrada por AG con menor número de carbono (grasa más blanda).
El punto de fusión determina si el lípido es líquido o sólido a temperaturas ambiente.
El punto de fusión depende principalmente del grado de saturación y en menor grado por la longitud de la cadena carbonada. En general los que tienen menos de diez carbonos son líquidos.
Los AG de cadena larga tienen entre 4 y 24 átomos de carbono y un solo grupo carboxilo, predominando los de cadena entre 14 y 18 carbonos (Tabla 1).
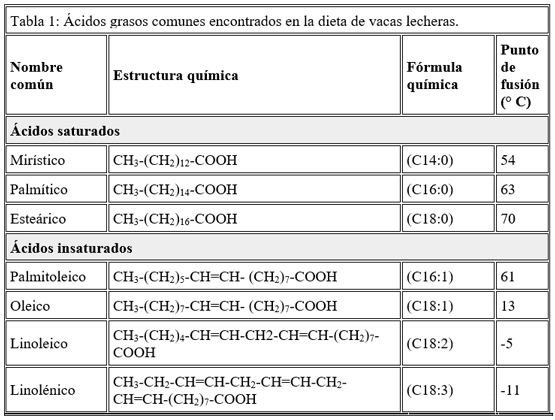
Las grasasvegetales tienen entre el 70 a 80% de AG insaturados y tienden a quedarse en estado líquido (denominado comúnmente aceites). Por otro lado, las grasas de origen animal contienen 40-50% de AG saturados y tienden a quedarse en estado sólido (denominado comúnmente grasas). El grado de saturación tiene un efecto marcado en la digestión y, en el caso de los rumiantes, puede interferir con la fermentación de la fibra en el rumen.
B. Ácidos grasos esenciales.
Difícilmente se dan carencias o enfermedades carenciales por ausencia de AG en la ración, pero en circunstancias muy especiales o experimentalmente se pueden citar problemas de crecimiento y dermatológicos en ratas con dietas carentes de grasa. Los AGesenciales son el linoleico, linolénico y araquidónico.
C. Ceras
Son lípidos sencillos compuestos por un ácido graso de cadena larga esterificado con un alcohol de alto peso molecular. Carecen de valor nutritivo puesto que no son digeridas por animales. Suelen constituir las cubiertas protectoras de tejidos animales (lana, plumas) y vegetales.
2. Lípidos compuestos saponificables.
Son ésteres de los ácidos grasos con grupos polares hidrófilos (bases nitrogenadas, azúcares) además de los restos de ácidos graso hidrófobos.
A. Fosfolípidos
Esteres del glicerol en el que dos grupos hidroxilo se esterifican con 2 AG de cadena larga y 1 grupo con ácido fosfórico. Presentan propiedades emulsionantes y realizan importantes funciones en el transporte de lípidos en la sangre.
Los más abundantes en animales y vegetales son las lecitinas en las que el ácido fosfórico se encuentra también esterificado con la base nitrogenada colina.
Los fosfolípidos son componentes menores en los alimentos, encontrados principalmente en las bacterias del rumen. Abundan en los forrajes.
B. Glicolípidos
Los glicolípidos son una segunda clase de lípidos que se encuentran principalmente en los forrajes (gramíneas y leguminosas).
Tienen una estructura parecida a los triglicéridos con la excepción que uno de los tres AG ha sido sustituido por un azúcar (usualmente galactosa). Dos grupos hidroxilo del glicerol se esterifican con AG especialmente linoleico. Al otro grupo hidroxilo se incorporan una o dos moléculas de galactosa.
C. Esfingolípidos
Contienen el amino-alcohol esfingosina en vez del glicerol, al que se le añaden un ácido graso, fosfato y colina. Abundan en las membranas del tejido nervioso.
D. Lipoproteínas
Son lípidos asociados a proteínas específicas. Importantes en el transporte de lípidos por vía sanguínea.
Lípidos no saponificables.
No contienen ácidos grasos y no pueden formar jabones.
A. Esteroides
Grupo de compuestos fisiológicamente importantes en animales y vegetales derivados del ciclopentanoperhidrofenantreno.
B. Terpenos.
Tienen olores y sabores característicos. Al degradarse producen isopreno. No proporcionan energía a los animales.
Dentro de este grupo también se incluyen los carotenoides, pigmentos vegetales y vitaminas liposolubles, como lípidos no saponificables y misiones específicas en el interior del organismo.
Digestión y absorción de grasas.
El objetivo primario de la digestión de los lípidos es hacerlos hidromiscibles y puedan absorberse a través de las microvellosidades intestinales que están recubiertas por una capa acuosa.
Hidrólisis y saturación de lípidos en el rumen
En el rumen, la mayoría de los lípidos son hidrolizados. Se rompe el enlace entre el glicerol y los tres ácidos grasos. El glicerol se fermenta rápidamente para formar AGV. Algunos ácidos grasos son utilizados por las bacterias para sintetizar los fosfolípidos necesarios para construir sus membranas de células.
Otra acción importante de los microorganismos (MO)del rumen es de hidrogenar los AG insaturados. En este proceso, un ácido graso resulta saturado porque un enlace doble se reemplaza por dos átomos de hidrogeno. Por ejemplo, la hidrogenación convierte el ácido oleico en ácido esteárico.
Los AG libres (AGL) en el rumen tienden a ligarse a partículas de alimentos y MO y propiciar más fermentaciones, especialmente de los carbohidratos fibrosos. La mayoría de los lípidos que salen del rumen son AG saturados (85-90%) principalmente en la forma de ácidos palmítico y esteárico ligados a partículas de alimentos y MO y los fosfolípidos microbianos (10-15%).
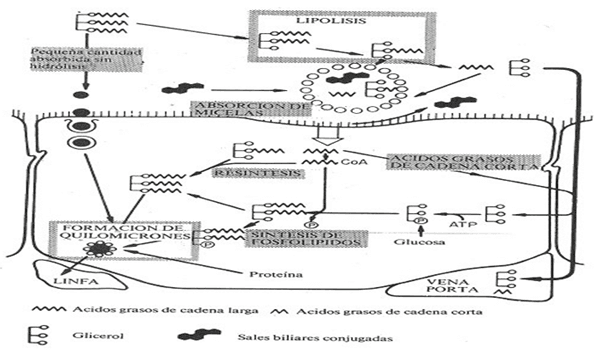
Absorción intestinal de los lípidos
Los fosfolípidos microbianos y los ácidos grasos procesados son digeridos y absorbidos a través de la pared del intestino. La bilis secretada por el hígado y las secreciones pancreáticas (ricas en enzimas y en especial las lipasas pancreáticas y bicarbonato) se mezclan con el contenido del intestino delgado. Las secreciones biliares en especial los ácidos glicocólico, taurocólico y cólico son esenciales para preparar los lípidos para absorción, formando partículas mezclables con agua que pueden entrar en las células intestinales. En las células intestinales la mayor parte de los ácidos grasos se ligan con glicerol (proveniente de la glucosa de la sangre) para formar triglicéridos.
Los triglicéridos, algunos AGL, colesterol y otras sustancias relacionadas con lípidos se recubren con proteínas para formar lipoproteínas ricas en triglicéridos, también llamados lipoproteínas de baja densidad (LDL). Las lipoproteínas ricas en triglicéridos entran en los vasos linfáticos y de allí pasan al canal torácico (donde el sistema linfático se conecta con la sangre) y así llegan a la sangre. En contraste con la mayoría de nutrientes absorbidos en el tracto gastrointestinal los lípidos absorbidos no van al hígado, sino que entran directamente a la circulación general. Así los lípidos absorbidos pueden ser utilizados por todos los tejidos del cuerpo sin ser procesados por el hígado.
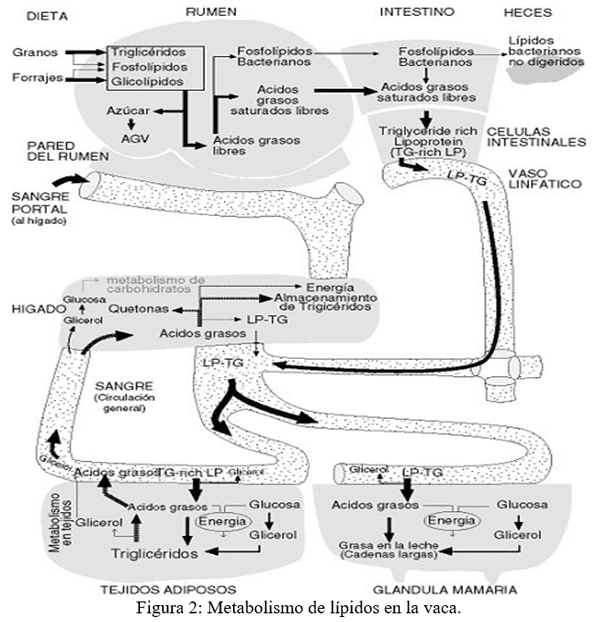
Utilización de los lípidos de la ración
Casi la mitad de la grasa presente en la leche deriva del metabolismo de lípidos en la glándula mamaria. Estos ácidos grasos provienen principalmente de las lipoproteínas ricas en triglicéridos. Un aumento de AG con más de 16 átomos de carbono en la dieta aumenta su secreción en la leche, pero también inhibe la síntesis de ácidos grasos de cadena corta y mediana.
Así la depresión marcada en la secreción de grasa en la leche cuando se alimenta las vacas con dietas bajas en fibra no puede ser compensando dando más grasa en la dieta.
Catabolismo de los lípidos
El principal mecanismo de obtención de energía de los lípidos lo constituye la oxidación de los ácidos grasos, que se obtienen de los triglicéridos mediante hidrólisis por lipasas específicas. Éstos siempre podrán entrar en el ciclo de Krebs, por lo que cuanto más largo sea el ácido graso mayor cantidad de energía se obtendrá en su oxidación. La glicerina también podrá degradarse si se transforma en dihidroxiacetona, entrando en la glucólisis.
Oxidación de los ácidos grasos
Los excesos de glúcidos ingeridos en la dieta se almacenan, para ser movilizados cuando el organismo lo necesite durante los periodos de ayuno o de demanda energética excesiva. Los triglicéridosson especialmente aptos para esta función. El principal mecanismo de obtención de energía de los lípidos lo constituye la llamada beta-oxidación de los ácidos grasos.
Los ácidos grasos que no se utilizan para sintetizar eicosanoides ni se incorporan a los tejidos se oxidan para producir energía.
Los AG proporcionan energía a través de la beta oxidación que tiene lugar en las mitocondrias de todas las células, excepto las del cerebro y las del riñon. Entran en la mitocondria en forma de los correspondientes derivados de acil carnitina. Los AG saturados de cadena corta, media y larga se someten al primer paso de la beta oxidación con distintas deshidrogenasas.
El proceso va generando sucesivamente moléculas de acetil-CoA que entran en el ciclo de los ácidos tricarboxílicos o en otras rutas metabólicas.
El acetil-coenzima A se incorpora al ciclo de Krebs para continuar su degradación. Como ejemplo el balance energético de 1 mol de ácido palmítico (16 átomos de carbono) da lugar a 129 ATP.
El producto final de los AG con un número par de átomos de carbono es el acetato.
Los AG insaturados requieren dos pasos enzimáticos más que los AG saturados para cambiar los dobles enlaces en cis a trans y para desplazarlos de la posición alfa a la beta.
Aun así, la oxidación de los AG insaturados, incluidos el ácido linoleico, es tan rápida o más que la del ácido palmítico. La reacción de la oxidación inicial es realizada por una enzima distinta de la que se encuentra en las mitocondrias; el acil-CoA graso entra directamente en esta organela. El proceso no conduce a la producción completa de acetato, sino que a la mitocondria se transfiere un ácido graso acortado para completar la oxidación.
Los AG de cadena larga (>20C) son oxidados preferentemente por los peroxisomas; también los ácidos grasos con menos de 14C se oxidan mediante este sistema.
La oxidación peroxisomal es normalmente menos eficaz que la mitocondrial y produce más calor. Este tipo de oxidación puede inducirse con alimentos de alto contenido en grasas.
El papel de hígado en la movilización de lípidos
En periodos de sub-alimentación o en la primera parte de lactancia, las vacas suplen su demanda energética movilizando los tejidos adiposos ya que la energía proveniente de la dieta no es suficiente. Los ácidos grasos de los triglicéridos almacenados en los tejidos adiposos (ubicados principalmente en el abdomen y encima de los riñones) son liberados hacia la sangre. Los AG liberados son absorbidos por el hígado donde pueden ser utilizados como fuente de energía o pueden ser liberados hacia la sangre y utilizados como una fuente de energía en muchos tejidos. El hígado no tiene una alta capacidad para formar y exportar lipoproteínas ricas en triglicéridos y los AG excedentes son almacenados como triglicéridos en el hígado.
La grasa depositada en el hígado hace difícil al hígado formar más glucosa. Esta condición ocurre principalmente en los primeros días de lactancia y puede llevar a desordenes metabólicos como cetosis e hígado graso.
Hígado graso
El hígado graso es un síntoma de un trastorno del metabolismo de las grasas relacionado con la sobreproducción de grasa en el hígado y un mayor transporte de grasa en el hígado o bien por la subutilización de grasa en el hígado o una defectuosa liberación de las mismas.
Colesterol
El colesterol se encuentra en todos los tejidos animales, es componente esencial de las membranas además de ser precursor de sustancias muy útiles al organismo como hormonas esteroides, ácidos biliares o vitamina D. Sin embargo, factores exógenos como raciones ricas en grasas o con elevada proporción de ácidos graso saturados, hacen que el contenido en colesterol del organismo y del plasma se eleve con el consiguiente riesgo de aparición de trastornos vasculares y cardíacos.
Universidad de Córdoba, España. https://www.uco.es/zootecniaygestion/menu.php?tema=154
Temas relacionados:
Autores:


Recomendar
Comentar
Compartir

Orffa Excentials
10 de abril de 2021
Estimados colegas,
¿Tienen experiencia con emulsificantes?
Recomendar
Responder


7 de julio de 2020
Sí, en efecto sobre grasas en la alimentación animal se han escrito libros de 500 páginas. Hay un mundo de información. Detallarlo en un foro no alcanza el tiempo y espacio.
No hay 2 aceites iguales cartamo, linaza, ajonjolí, soya, girasol, canola, maíz, etc. No hay 2 grasas iguales ovinos, res, leche, cerdo, pollos, pescado, etc.
En corral de engorda la carne mas sabrosa se obtiene sin la inclusión de grasas adicionales en la dieta. Se aplican un poco de aceite 3% o menos para reducir el polvo en la planta de alimentos balanceados y no como recurso energético. Es un pre vehículo para las VITAMINAS liposolubles y asegurar la presencia de aceites grasos esenciales. Solo por esos fines se incluyen, nada mas. No por su valor energético. ESO SE VALORA EN LAS DIETAS PORCINAS, AVES, CAMARONES, ACUICULTURA y mascotas.
Para mejorar la calidad y sabor de la carne de res no se incluyen aceites y grasas.
Recomendar
Responder

7 de agosto de 2020
De qué manera opcional y natural podemos brindar un alimento equilibrado a los rumiantes para adquirir peso y y excelente respuesta con calidad
Recomendar
Responder

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Usuarios destacados en Lechería






Special Nutrients
Special Nutrients de Estados Unidos produce adsorbentes de micotoxinas para la alimentación animal y la agricultura orgánica mundial

Priya Chemicals
Priya Chemicals es el mayor fabricante de aminoácidos y productos a base de aminoácidos para el uso en el ámbito de nutracéuticos, Agricultura y Veterinaria