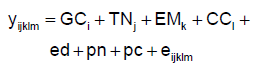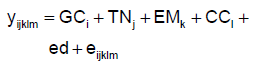INTRODUCCIÓN
Las parasitosis gastrointestinales (PGI) causadas por nematodos, constituyen una de las principales limitantes sanitario-económicas para la producción ovina en el Uruguay y en el mundo (Castells et al., 1995, Pandey, 1999; Perry y Randolph, 1999; Nieto et al., 2002). El incremento de la resistencia de los PGI a las drogas antihelmínticas (ATH) ha sido descrito en varios países del mundo, incluyendo Uruguay, donde la problemática se ha venido agravando en los últimos años (Nari et al., 1996; Waller, 1997). Esto ha determinado un cambio de enfoque en el control de los PGI, donde si bien los ATH siguen ocupando un lugar preponderante, otros métodos de control deben ser incorporados apuntando a lo que se denomina Control Integrado de Parásitos (Castells, 2002).
Una de las estrategias propuestas que se manejan es la selección de animales genéticamente resistentes a los PGI, utilizando como criterio, el recuento de huevos por gramo de materia fecal (HPG). Estos animales presentan una menor carga parasitaria, lo que contribuye a reducir la contaminación de las pasturas así como los requerimientos en el uso de ATH; retardando así el desarrollo de la resistencia y disminuyendo los costos del tratamiento (Nieto et al., 2002; Bishop y Stear, 2003).
Actualmente en el Uruguay, se realiza la evaluación genética de la resistencia genética a PGI en las razas Merino y Corriedale, publicándose el valor genético como diferencia esperada en la progenie (DEP) del recuento de HPG.
A partir del año 1998, el Instituto Nacional de Investigación Agropecuaria (INIA), el Secretariado Uruguayo de la Lana (SUL) y la Sociedad de Criadores Merino Australiano del Uruguay (SCMAU), comenzaron con la ejecución de un proyecto de investigación y desarrollo del Merino Fino para Uruguay. Un componente fundamental en el proceso de formación de dicho Proyecto, fue la formación del Núcleo Fundacional de Merino Fino de la Unidad Experimental “Glencoe” de INIA Tacuarembó (NFG). EL NFG fue formado por 475 borregas provenientes de 36 productores colaboradores, las que fueron inseminadas con semen perteneciente a carneros importados y nacionales dando lugar, en 1999, a la primera progenie del NFG. Las estrategias empleadas para la creación de este Núcleo fueron detalladas por Montossi et al. (2005). Los animales pertenecientes al NFG, al nacer dentro del sistema de evaluación, cuentan con DEP de HPG de borregos.
Según varios autores (e.g., Pandey, 1999; McClure, 2000; Vanimisseti, 2003) los principales factores que influyen en las variaciones que se observan en cuanto a la resistencia a PGI entre los animales incluyen: raza, individuo, sexo, edad, estatus fisiológico y reproductivo del animal (i.e. preñez, lactancia), tipo de parto, nutrición, y factores estresantes como el destete, el clima y el manejo.
El objetivo del presente trabajo es determinar qué factores de origen no genético afectan el recuento de HPG en ovinos de la raza Merino del Uruguay pertenecientes a un núcleo de selección.
MATERIALES Y MÉTODOS
Los animales empleados en el presente estudio pertenecen al NFG, de INIA Tacuarembó. La recolección de muestras de materia fecal se realizó según el protocolo establecido para las evaluaciones genéticas de Uruguay (Castells, 2009). Los registros comienzan a partir del destete, con 2 muestreos realizados en ciclos parasitarios independientes y bajo infección natural. En el momento del destete (aproximadamente a los cuatro meses de edad), se dosifica a todos los corderos, y a los 10 días se chequea que la carga parasitaria sea menor de 100 HPG. A partir de ese momento, se realizan monitoreos cada 15 días sobre 20 corderos elegidos al azar, hasta que la media aritmética supera los 500 HPG y no más de 20% de la muestra presenta conteos menores a 100. En ese momento, se realiza el primer muestreo general (HPG1), donde se sacan muestras individuales de todos los corderos para ser analizadas en el laboratorio. Una vez procesadas las mismas, los corderos nuevamente son dosificados, a los 10 días se chequea, luego se monitorea y cuando nuevamente se superan los 500 HPG se realiza el segundo muestreo general (HPG2). Una vez que se terminan de procesar las muestras en el laboratorio, los corderos se dosifican por tercera vez. Para el recuento de HPG la técnica empleada es la de Mc Master modificada (Withlock, 1948), con una sensibilidad de 100.
Debido a que la variable HPG no presenta una distribución normal, es necesaria la transformación de los datos para llevar a cabo pruebas estadísticas paramétricas. La variable analizada es la resultante del logaritmo natural del HPG más 100 (LnHPG) como se detalla en Ciappesoni et al. (2010).
El análisis estadístico se realizó mediante el procedimiento GLM del paquete estadístico SAS (SAS, 2003), para el cual se utilizaron 2.315 datos de HPG1 y 2.047 de HPG2, que fueron registrados entre los años 2001 al 2010. Se eliminaron aquellos registros donde el LnHPG 1 o 2 estuvo fuera del rango de ±3 desvíos estándar de la media del grupo año-sexo-lote de manejo correspondiente.
Asimismo, se analizaron que factores afectan al peso al nacimiento con el fin de esclarecer las relaciones entre los diferentes efectos y obtener los residuales para calcular las correlaciones fenotípicas con el HPG. Los modelos empleados para el análisis estadístico fueron los siguientes:
Modelo 1: HPG
Modelo 2: Peso al nacimiento
Donde: y es logaritmo natural de HPG (LnHPG 1 o 2) en el caso del modelo 1 y el peso al nacimiento en el modelo 2; GC, TN, EM y CC son los efectos fijos del grupo contemporáneo (año-sexo-lote de nacimiento-lote de destete-época de nacimiento) (76 y 71 niveles para LnHPG1 y LnHPG2 respectivamente), tipo de nacimiento (2 niveles), edad de la madre (4 niveles) y condición corporal de la madre al parto (3 clases: ≤3, >3 y ≤4, y >4) respectivamente; ed es la edad del cordero en días a la medición, pn es el peso al nacimiento (kg), pc es el peso corporal más próximo a la medición (peso al destete para el LnHPG1 y peso a la esquila para el LnHPG2, en kg), todos ellos como covariables; y e es el residuo aleatorio.
El efecto época de nacimiento se definió según la fecha de nacimiento formando grupos cada 15 días desde que nace el primer cordero en el año. Se obtuvieron cuatro niveles, ya que ningún nacimiento se extendió más allá de 60 días desde el primer nacimiento.
Con respecto al GC, es importante destacar que los lotes de nacimiento se realizaron sólo en los años 2005 y 2008 en base a la fecha de parto promedio esperada (sin tener en cuenta el tipo de nacimiento), y sólo se hicieron lotes de destete entre los años 2007 y 2009. Por otro lado, el GC para el LnHPG1 tuvo más niveles que el LnHPG2, debido a que en el año 2008, las hembras no alcanzaron a tener registros de HPG2; ya que el recuento de huevos no se elevó lo suficiente hasta pasados los 500 días de edad en promedio.
Los residuales del modelo 1 para ambas características (LnHPG 1 y 2) fueron analizados con el procedimiento UNIVARIATE (SAS, 2003). La normalidad de LnHPG1 y LnHPG2 fue analizada mediante gráficas tipo PP (percentil-percentil) de ajuste a la distribución e histogramas, y se realizaron los test de hipótesis de normalidad de Kolmogorov-Smirnov, Cramér-von Mises y Anderson-Darling. Asimismo, estos residuales y los del modelo 2 (pn) fueron utilizados para calcular las correlaciones fenotípicas entre las características (pn, LnHPG1 y 2) con el procedimiento CORR (SAS, 2003).
RESULTADOS Y DISCUSIÓN
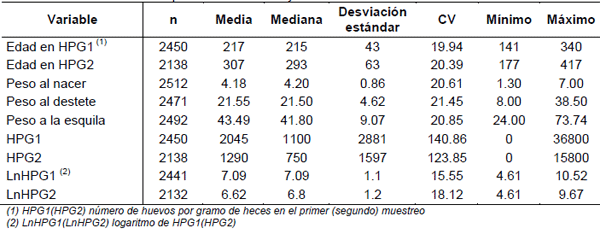
En el Cuadro 1 se muestra la estadística descriptiva de los efectos y características analizadas. La amplia variación observada en las edades de registro del HPG 1 y 2 es debida a que se trabaja con infecciones naturales, dependiéndose de la epidemiología de la parasitosis en cada año.
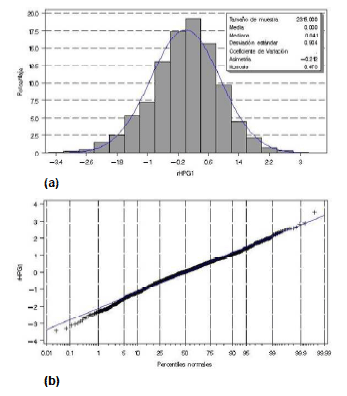
Los resultados para los registros de HPG 1 y 2 están dentro de lo esperado para el tipo de distribución no normal descrita. Adicionalmente, la moda para HPG 1 y 2 fue de 200 y 0 HPG respectivamente. Estos valores fueron similares a los obtenidos por Castells (2009) donde la media, la mediana y la moda no coinciden. En la figura 1 se encuentra representado gráficamente los residuales del modelo 1 para LnHPG 1. En la figura 1a se observa una distribución que se asemeja a una campana de Gauss, y en la figura 1b los puntos están prácticamente alineados a la recta; por lo que los datos transformados (LnHPG 1) se distribuyen aproximadamente en forma normal. Resultados similares se obtuvieron para LnHPG 2. Contrariamente, los tres tests de normalidad presentaron valores de p<0.01, indicando que la distribución de los residuales analizados difiere de la distribución normal. Sin embargo, varios autores mencionan que para tamaños de muestras grandes, como en este estudio, cierto desvío de la normalidad puede ser tolerado respaldándose en el Teorema Central del Límite (Rice, 1995).
Figura 1: Distribución de los residuales del HPG1 transformados mediante el Logaritmo natural y agrupados en histogramas (a) y percentiles (b).
Cuadro 1. Estadística descriptiva de los efectos y características analizadas
Cuadro 2. Análisis de Varianza para el conteo del logaritmo de HPG (LnHPG 1 y 2)
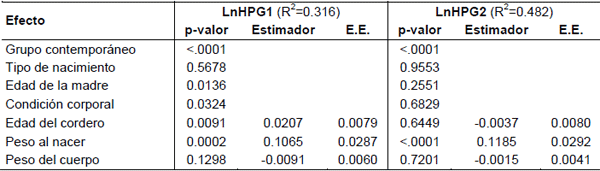
En el Cuadro 2 se observa el nivel de significancia estadística de los diferentes efectos sobre las variables LnHPG 1 y 2.
Para el LnHPG1, los efectos con significancia estadística (p<0.05) fueron la condición corporal y edad de la madre al parto (CC y EM), el peso del cordero al nacimiento (pn), la edad del cordero al momento de la medición y el grupo contemporáneo (GC).
La CC tuvo un efecto positivo sobre el HPG, es decir, los corderos de madres de alta CC, tuvieron menor HPG (ver Cuadro 3). Si la mediana de HPG de la población fuese 1000, los corderos de madres de CC baja, eliminarían 208 huevos más con respecto a los corderos cuyas madres son de CC alta. Esto puede deberse a que las madres con mejor CC tendrán una mayor producción de leche, por lo que los corderos estarán mejor nutridos, siendo menos susceptibles a las infecciones parasitarias. Banchero et al. (2005) observaron que corderos nacidos de ovejas de alta CC tenían mayor vigor y mamaban más del doble que los corderos nacidos de ovejas de baja CC. Según Iposu (2007), el amamantamiento podría aumentar la nutrición proteica de los corderos y consecuentemente acelerar la adquisición de la inmunidad, particularmente cuando se exponen a las larvas de los nematodos en etapas tempranas de la vida. Por otro lado, el aumento en la demanda nutricional de la hembra durante la progresión de la gestación y en la lactación subsiguiente, en tiempos de escasez de nutrientes, resulta en una penalidad sobre la expresión de la inmunidad adquirida a los parásitos, y por lo tanto en una mayor eliminación de huevos (Houdijk, 2008). De esta manera, si la CC es buena, habrá un menor estrés nutricional de la madre, pudiéndose reflejar en un alza de lactación menos pronunciada. En concordancia con este último punto, Hayward et al. (2010) encontraron una asociación negativa entre el HPG de la oveja y su condición corporal, es decir, aquellas con menor CC eliminarían más huevos durante la lactancia.
Cuadro 3. Medias de mínimos cuadrados para el LnHPG1 según la condición corporal (CC) y edad de la madre (EM)

Con respecto a la EM, los corderos de madres de mayor edad presentaron conteos de HPG menores (ver Cuadro 3). En una población cuya mediana de HPG fuese 1000, las hembras de la clases 1, 2 y 3 eliminarían 157, 166 y 146 huevos más, respectivamente, con respecto a las hembras de la clase 4. Esto podría verse explicado por el hecho de que, ovejas de mayor edad producen mayor cantidad de leche y por otro lado, Watson et al. (1995), Pandey (1999) y Bishop y Stear (2001), encontraron que a medida que aumenta la edad de la oveja disminuye el HPG en el período del periparto; disminuyendo de esta manera la contaminación de las pasturas. Sin embargo, Woolaston (1992) y Morris et al. (1998) no encontraron variaciones significantes asociadas a la edad. Hayward et al. (2010) encontraron que corderos nacidos de ovejas de edad media, tenían menor HPG que aquellos nacidos de ovejas jóvenes o de edad avanzada. A su vez, observaron en hembras adultas, una asociación negativa entre el HPG y la edad en que comenzaban a reproducirse, sugiriendo que las ovejas que comienzan en su primer año, tienen mayor HPG, que aquellas que lo hacen en el segundo y tercer año de vida.
La edad del cordero a la medición también tuvo un efecto significativo, teniendo los recuentos más elevados los corderos de mayor edad. Posiblemente se deba al hecho de que, éstos animales estuvieron más tiempo en las pasturas, y consecuentemente, tuvieron una mayor exposición a las larvas infestantes.
Para el LnHPG2, los efectos con significancia estadística fueron el pn y el GC. El coeficiente de determinación (R2) fue mayor que para el LnHPG1 (0.32 vs 0.48). Seguramente la no significancia de los efectos maternales (CC y EM) se deba a que este segundo conteo se realizó en promedio a los 10 meses de edad, siendo muy lejano del destete (a los cuatro meses en promedio). Asimismo, el efecto de la edad a la medición no fue significativo porque los animales dentro del mismo GC (que corrige a su vez por época de nacimiento) cuentan con la misma cantidad de días desde el tratamiento antihelmíntico anterior. El pn tuvo un efecto positivo para ambos recuentos, es decir, un mayor HPG en corderos más pesados al nacimiento. El efecto de aumentar un kg de peso al nacer se correspondería con el aumento de 100 HPG aproximadamente, en una población con una mediana de 1000 HPG. También se observó el mismo efecto (p<0.05), al sustituir en el modelo 1 el pn como covariable por un efecto fijo de tres clases de pn (≤3.5, 3.5-5.0 y >5.0 kg) (los resultados no se muestran), presentando los conteos más bajos los corderos del primer nivel. En una población con una mediana de 1000 HPG, los corderos del primer nivel, eliminarían 200 huevos menos con respecto a los del tercer nivel, tanto para el LnHPG1 como para el LnHPG2. Esto puede deberse a efectos del propio cordero y a un efecto ambiental de la madre. Bishop y Stear (2001) encontraron una correlación genética baja pero desfavorable entre el HPG de la oveja en el periparto con el pn del cordero y con el peso promedio del cordero a las 4 semanas de edad (0.11±0.10 y 0.24±0.10 respectivamente). Estimas similares fueron reportadas por el Instituto Roslin (2000), donde la correlación genética entre el HPG de la oveja con el peso del cordero al nacimiento, a las 4 y a las 8 semanas de edad, fueron de 0.19±0.11, 0.24±0.10 y 0.20±0.11, respectivamente. Por lo tanto, aquellos vientres con corderos más pesados, tendrían mayor HPG, aumentando a su vez la oferta larvaria en las pasturas y la probabilidad de infección de los corderos recién nacidos. Es así que Morris et al. (1998), estimaron una regresión fenotípica entre el HPG de la oveja y el de su/s cordero/s en la misma lactación de 0.29±0.06. Se postula que habría una competencia entre el esfuerzo puesto en criar al cordero y en el de controlar la propia infección parasitaria (Bishop y Stear, 2001). Asimismo, Vanimisseti (2003) establece que, la producción de corderos más pesados puede aumentar el estrés de la madre durante la lactación, y puede llevarle más tiempo para recuperarse de los efectos de la lactancia, lo que deriva en una mayor susceptibilidad a la infección parasitaria. Este mismo autor encontró que ovejas con un mayor mérito genético para crecimiento como corderas: 1) fueron menos resistentes a las infecciones como adultos, con mayor conteo de HPG pos-infección; aunque las ovejas que eran fenotípicamente más pesadas en el momento de la infección tuvieron mayor capacidad para enfrentar a la misma indicado por el mayor hematocrito, y 2) produjeron corderos con mayor potencial de crecimiento y por lo tanto más demandantes en cuanto a la necesidad de nutrientes. Por otra parte, Dwyer et al. (2001), trabajando con dos líneas divergentes seleccionadas por el contenido de grasa en la canal (sin afectar el peso vivo a la faena) de ovinos Scottish Blackface, observaron que el pn en los corderos de la línea magra fue menor, siendo éstos más activos, más rápidos en pararse y en buscar la ubre y mamar, que aquellos de la línea grasa. Las diferencias entre ambas líneas podría repercutir en una mejor respuesta inmunitaria, pudiendo explicar en parte, por qué los corderos más livianos tienen menor recuento de HPG.
Cuadro 4. Medias de mínimos cuadrados para el peso al nacimiento (kg) según el tipo de nacimiento (TN), la condición corporal (CC) y edad de la madre (EM)
Todos los efectos incluidos en el modelo 2 fueron significativos (p<0.0001) para el peso al nacimiento. Las medias de cuadrados mínimos para los efectos en el modelo 2 se presentan en el Cuadro 4. La correlación entre los residuales del peso al nacimiento con los residuales de LnHPG 1 y 2 fue positiva y de baja magnitud (0.075 y 0.088 respectivamente). Estos valores son ligeramente inferiores a los estimadores del efecto del pn presentados en el Cuadro 3 para ambas mediciones (LnHPG 1 y 2). Si bien ambas correlaciones son diferentes de cero (p=0.0078 y p<0.0001, respectivamente) el aumento del HPG al aumentar un kg del pn (i.e., 80 HPG en una población con mediana de 1000 HPG) es poco relevante.
Cuando se analizó el efecto época de nacimiento-año sobre el HPG, se observó que no hubo una consistencia entre un año y otro, debido a las diferentes condiciones climáticas entre los distintos años. En determinados años, puede suceder que los corderos que nacen al principio del período de parición tengan los menores recuentos de HPG de borregos, mientras que en otros años, frente a otras condiciones ambientales, los corderos con menor HPG sean los que nazcan hacia el final del período.
En concordancia con Bouix et al. (1998), el tipo de nacimiento no fue significativo para ninguna de las dos variables. Sin embargo, Hayward et al. (2010) encontraron que los corderos nacidos como mellizos, exhibieron mayor HPG que los nacidos como únicos. Diversas causas pueden explicar este resultado. En primer lugar, los mellizos serán más susceptibles a las infecciones parasitarias, ya que generalmente nacen con menor peso y en peores condiciones. En nuestro modelo esto se estaría reflejando en el efecto del pn. Por otro lado, teniendo en cuenta que el amamantamiento prolongado provee una mayor resistencia parasitaria en corderos (posiblemente debido a un efecto positivo sobre la ganancia de peso), los mellizos reciben menor cantidad de nutrientes de la madre, lo que los hace más susceptibles. Adicionalmente, un mayor esfuerzo reproductivo de las madres con gestaciones múltiples, se refleja en un alza de lactación más pronunciada lo que da lugar a un ambiente contaminado para los recién nacidos. Diversos estudios describen que las ovejas con mellizos tienen mayor conteo de huevos que las que tienen un único cordero (Woolaston, 1992; Watson et al., 1995; Morris et al., 1998; Pandey, 1999; Bishop y Stear, 2001). Asimismo, el tipo de nacimiento está relacionado con la condición corporal de la madre. En los datos analizados, el 36.3% de las madres con gestación doble o triple fueron valoradas con condición corporal menor o igual a tres contra sólo un 16.9% de las madres con gestación simple. Adicionalmente, sólo un 8.4% de las melliceras o trilliceras obtuvieron una valoración mayor a cuatro contra un 25.4% de aquellas con un único cordero.
La correlación fenotípica entre LnHPG 1 y 2 fue 0.18 (p<0.0001), valor similar al obtenido en estudios realizados en el total de la población evaluada genéticamente de la raza Merino (Ciappesoni, comunicación personal).
CONCLUSIONES
Los efectos maternales (condición corporal y edad de la madre al parto) presentan una mayor incidencia sobre el primer conteo de HPG (LnHPG1) mientras que para el LnHPG2 éstos dejan de ser significativos. El peso al nacimiento presenta un efecto en ambos conteos de HPG, incluso cuando se corrige por los factores de tipo de nacimiento, la edad de la madre y su condición corporal al parto, sin embargo este efecto es de poca relevancia.
Pese a presentar efectos significativos, la CC y el pn no son incluidos como efectos en las evaluaciones genéticas rutinarias. Esto se debe a que no se registran en todas las cabañas. Además, el pn contiene un componente genético lo que provocaría que su inclusión en el modelo afecte la evaluación del HPG en sí mismo.
A su vez, dado que las características y los efectos analizados se encuentran estrechamente relacionados, es importante distinguir entre los conceptos significancia estadística y relevancia, y si la asociación encontrada presenta una explicación biológica real aparente.
AGRADECIMIENTOS
A INIA por la concesión de una beca de inicio en la investigación a la Dra. Virginia Goldberg y por la financiación del presente trabajo.
Al personal de Glencoe por la recolección de las muestras y al Laboratorio de Sanidad de INIA Tacuarembó donde se llevó a cabo el procesamiento de las mismas.
REFERENCIAS BIBLIOGRÁFICAS
BANCHERO, G., QUINTANS, G., MILTON, J. T. B. y LINDSAY, D. 2005. Comportamiento maternal y vigor de los corderos al parto: efecto de la carga fetal y la condición corporal. En: Seminario de actualización técnica: Reproducción Ovina. Recientes avances realizados por INIA. SAD N° 401: 61-67.
BISHOP, S. C. y STEAR, M. J. 2001. Inheritance of faecal egg counts during early lactation in Scottish Blackface ewes facing mixed, natural nematode infections. Anim. Sci. 73: 389-395.
BISHOP, S. C y STEAR, M. J. 2003. Modeling of host genetics and resistance to infectious diseases: understanding and controlling nematode infections. Vet. Parasitol. 115: 147-166.
BOUIX, J., KRUPINSKI, J., RZEPECKI, R., NOWOSAD, B., SKRZYZALA, I., ROBORZYNSKI, M., FUDALEWICZ-NIEMCZYK, W., SKALSKA, M., MALCZEWSKI, A. Y GRUNER, L. 1998. Genetic resistance to gastrointestinal nematode parasites in Polish longwool sheep. Int. J. Parasit. 28: 1797-1804.
CASTELLS, D., NARI, A.; RIZZO, E.; MARMOL, E. y ACOSTA, D. 1995. Efectos de los nematodos gastrointestinales sobre diversos parámetros productivos del ovino en la etapa de recría. Producción Ovina. 8: 17-32.
CASTELLS, D. 2002. Nuevo enfoque en el control parasitario de ovinos. En: Resistencia genética del ovino y su aplicación en sistemas de control integrado de parásitos; FAO Animal Production and Health Paper. Castells, D. FAO, Roma (Italia). Dirección de Producción y Sanidad Animal, pp. 17-24.
CASTELLS, D. 2009. Evaluación de resistencia genética de ovinos Corriedale a los nematodos gastrointestinales en Uruguay: Heredabilidad y correlaciones genéticas entre el recuento de huevos de nematodos y características productivas. Tesis de Maestría en Producción Animal. Universidad de la República, Montevideo, Uruguay. 40p.
CIAPPESONI, G., GIMENO, D. y RAVAGNOLO, O. 2010. Genetic Relationships between Faecal Worm Egg Count and Production Traits in Merino Sheep of Uruguay. Proc 9th WCGALP. Leipzig, Germany.
DWYER, C. M., LAWRENCE, A. B. y BISHOP, S. C. 2001. The effects of selection for lean tissue content on maternal and neonatal lamb behaviours in Scottish Blackface sheep. Anim. Sci. 72: 555-571.
HAYWARD, A. D., PILKINGTON, J. G., PEMBERTON, J. M. y KRUUK, L. E. B. 2010. Maternal effects and early-life performance are associated with parasite resistance across life in free-living Soay sheep. Parasitol. 1-13. HOUDIJK, J. G. M. 2008. Influence of periparturient nutritional demand on resistance to parasites in livestock. Parasite Immunology. 30: 113-121.
INSTITUTO ROSLIN. 2000. Genetic control of resistance to gastrointestinal parasites in hill sheep. Final Report. 11p. IPOSU, S. O. 2007. Effect of suckling to nematode parasites in young lambs. Thesis for the degree of Doctor of Philosophy. Lincoln University, Canterbury, New Zealand. 174p.
McCLURE, S. 2000. Sheep immunity to gastrointestinal nematode parasite- Review 2000. CSIRO Mac master laboratory. Armidale, Australia, 14p.
MONTOSSI, F., DE BARBIERI, I., CIAPPESONI, G., RAVAGNOLO, O., DE MATTOS, D., PÉREZ JONES, J., FROS, A., GRATTAROLA, M., MEDEROS, A. y SOARES DE LIMA, M. 2005. Núcleo fundacional de merino fino y superfino de la Unidad Experimental “Glencoe” - INIA Uruguay: una experiencia innovadora de mejoramiento genético asociativo y participativo. Agrociencia. 9 (1-2): 609-616.
MORRIS, C. A., BISSET, S. A., VLASSOFF, A., WEST, C. J. y WHEELER, M. 1998. Faecal nematode egg counts in lactating ewes from Romney flocks selectively bred for divergence in lamb faecal egg count. Anim. Sci. 67: 283-288.
NARI, A., SALLES, J., GIL, A., WALLER, P. J. y HANSEN, J. W. 1996. The prevalence of anthelmintic resistance in nematode parasites of sheep in Southern Latin America: Uruguay. Vet. Parasitol. 62: 213-222.
NIETO, L. M., MARTINS, E. N., MACEDO, F. A. F., SAKAGUTTI, E. S. y SANTOS, A. I. 2002. Genetic resistance to gastrointestinal endoparasites in different genetic groups of sheep. Proc 7th WCGALP, Montpellier, France.
PANDEY, V. S. 1999. Genetic resistance to internal parasites in sheep and goats and its exploitation for increasing animal productivity in Southeast Asia. Summary reports of European Commission supported STD-3 projects (1992-1995). pp.190-194.
PERRY, B. D. y RANDOLPH, T. F. 1999. Improving the assessment of the economic impact of parasitic diseases and of their control in production animals. Vet. Parasitol. 84: 145-168.
RICE, J. 1995. Mathematical Statistics and Data Analysis (Second ed.), Duxbury Press, ISBN 0-534- 20934-3. SAS. 2003.
SAS version 9.1.3 SAS Inst., Cary, N.C., USA.
VANIMISSETI, H. B. 2003. Genetics of resistance to Haemonchus contortus infections in sheep. Tesis de Master of Science, Blacksburg, Virginia. 73p.
WALLER, P. J. 1997. Anthelmintic resistance. Vet. Parasitol. 72: 391- 412.
WATSON, T. G., HOSKING, B. C., MORRIS, C. A. y HURFORD, A. P. 1995. Faecal nematode egg counts and haematology in Perendale ewes near lambing. Proc. N. Z. Soc. Anim. Prod. 55: 202-204.
WITHLOCK, H. V. 1948. Some modifications of the McMaster helminth egg counting technique and apparatus. J. Coun. Sci. Ind. Res. 21:177
WOOLASTON, R. R. 1992. Selection of Merino sheep for increased and decreased resistance to Haemonchus contortus: peri-parturient effects on faecal egg counts. Int. J. Parasit. 22 (7): 947-953