Introducción
Los forrajes provenientes de gramíneas y leguminosas representan el alimento más abundante para la vacas lecheras de México. Sin embargo, el valor nutricional de las especies forrajeras no siempre es el más apropiado para cubrir los requerimientos de los animales, especialmente durante la lactancia. Por ejemplo, la mayoría de las especies forrajeras tienen eficiencias de utilización baja debido a que la digestibilidad aparente de su fibra es inferior al 65.0 % (Van Soest, 1994). En el transcurso de los años, se han desarrollado procesos diferentes con fines de incrementar la digestibilidad y valor energético (Owens et al., 1997). Las acciones se hicieron con el objetivo de maximizar la producción de leche y reducir la incidencia de problemas metabólicos en el inicio de la lactancia, sin afectar los componentes de la leche, principalmente, su contenido graso y proteínico.
El tratamiento con enzimas fibrolíticas exógenas se propusó como un método para mejorar la calidad de los forrajes y el comportamiento animal. Sin embargo, los resultados obtenidos hasta ahora no son completamente satisfactorios. Varias enzimas, alimentos, animales, y factores de manejos están involucrados en la respuesta de los animales (Adesogan et al., 2014; Arriola et al., 2017).
Varios complejos enzimáticos se han propuesto para examinar la eficiencia de uso de las enzimas fibrolíticas exógenas en el mejoramiento de la producción de leche en vacas lecheras lactantes. En algunos estudios la producción de lehe incremento (Dyaa El-Din et al., 2013), y en otros no tuvo efectos (Refat et al., 2018). Tambien se observó mejoramientos en la digestibilidad de los forrajes (Chung et al., 2012; Phakachoed et al., 2013; Romero et al., 2015) cuando se suministró a dietas completas y en otros estudios se demostró lo contrario (Holsthausen et al., 2011). Además, se ha observado que tienen un efecto positivo en la eficiencia de utlización del alimento (Holsthausen et al., 2011; Refat et al., 2018).
Aunque se han publicado varios estudios de la actividad xilanásica en producción de leche, todavia no se define claramente, las dosis que pudieran ser apropiadas para mejorar dicho nivel sin afectar la composición de la mismas en el inicio de la lactancia. Con base en lo anterior, el objetivo del presente estudio fue determinar si la adición de 25 g animal-1 d-1 de un compuesto enzimático es apropiado para promover cambios en el consumo de materia seca, producción y composición de la leche de vacas Holstein-Frieisian que consumen dietas completas en condiciones de confinamiento.
Materiales y Métodos
Localización del experimento
El estudio se realizó en la lechería "18 de Julio", propiedad de la Universidad Autónoma Chapingo, ubicada en el municipio de Tlahualilo, Durango (25º 54'0'' N y 103 ° 35' 09'' W). La unidad de producción cuenta con aproximadamente 544 vacas en la línea de ordeño. El manejo de la alimentación en con base en dietas completas y ordeño mecánico. La altitud es de 1.137 m, el clima es desértico, la temperatura media anual es de 21.1 °C con 239 mm de precipitación anual, distribuidos principalmente de julio a septiembre (García, 1988).
Manejo y medición de las vacas
Los protocolos de investigación, el cuidado de los animales y los procedimientos de manejo se realizaron de acuerdo con las técnicas oficiales locales aprobadas de cuidado de los animales (NOM-051-ZOO-195 y NOM-024-ZOO-195). También se siguieron las técnicas y procedimientos de la Guía de Cuidado y Uso de Animales Agrícolas e Investigación y Enseñanza (Federación de Sociedades de Ciencia Animal, FASS, 2010). Los animales utilizados en el presente estudio fueron vacas gestantes de aproximadamente 20 días de su fecha probable de parto (con peso corporal (PV) de 780 ± 36.0 kg; de más de dos lactancias) . Las vacas seleccionadas fueron saludables con escore de condición corporal (ECC) de 3.3 ± 0.5 (escala: 1, muy delgada, a 5, muy gorda).
Al inicio del experimento, las vacas fueron estratificadas por PV y PL anterior; y se asignaron aleatoriamente a uno de dos tratamientos: 1) dieta completa (CONTROL, alfalfa-soya; n= 15); y 2) CONTROL + 25.00 g de una mezcla de enzimas fibrolíticas (Fibrozyme, Alltech, INC, Nichollsville, KY, USA; n=14). El compuesto enzimático es una combinación de extracto de fermentación de Aspergillus niger y Trichoderma viride, y fermentos solubles protegidos por técnicas de glucosilación. La actividad xilanásica es de 100 UI g-1; lo que significa que tiene el poder necesario de liberar 1.0 µmol de xilosa.
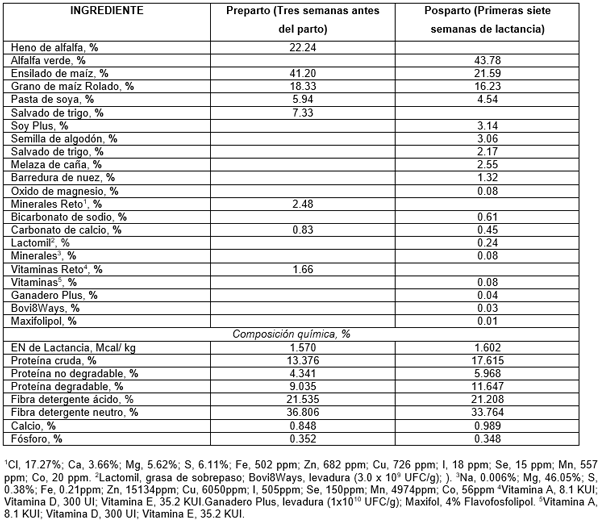
Las dietas se formularon siguiendo las recomendaciones del NRC (NRC, 2001) para vacas en período seco y lactancia. Los ingredientes, composición química y el valor nutritivo de las dietas experimentales se muestran en el Cuadro 1.
La alimentación se ofreció en tres partes iguales (33.33% de la dieta diaria total) a las 6:00, 14:00 y 22:00 h, lo cual es una práctica común en la industria lechera en está parte de México. Durante las primeras horas de la mañana, las vacas eligieron uno de los 30 alimentadores del tipo Calan Door (Calan Doors, Northwood, NH), de acuerdo con el tratamiento, cada vaca recibió la dieta correspondiente. La enzima se adicionó en el ofrecimiento de comida de la tarde (14:00 h), debido a que el tiempo disponible fue mayor en ese horario. Primero, se mezcló 25 g del complejo enzimático con 500 g de dieta y se ofreció a las vacas. Al terminar de consumir dicha porción se ofreció el resto del alimento correspondiente al horario.
Antes de cada tiempo de alimentación, el alimento ofrecido se pesó y se registró. Posteriormente, el alimento no consumido se pesó y se registró; este procedimiento se repitió cada vez que se ofreció alimento nuevo. Al final del día, las porciones no consumidas se sumaron y se restaron de la cantidad ofrecida para determinar el consumo de materia seca (CMS) individual diaria. El CMS se estableció proporcionando 15.0% más que el requerimiento de alimento sugerido por NRC (NRC, 2001), durante los primeros cuatro a seis días del período experimental.
El alimento consumido y el no consumido de cada vaca se muestrearon diariamente. Al final de la semana, las muestras de alimento consumido y no consumido se combinaron para formar muestras compuestas semanales. En ambos tipos de muestras, se determinó el contenido de materia seca (MS), proteína, fibra detergente neutro (FDN), fibra detergente ácido (FDA) y cenizas durante el período experimental.
Después de la obtención, las muestras se almacenaron a -20ºC. Posteriormente, se descongelaron, se pesaron y se secaron a 55.0 – 60.0°C en un horno de aire forzado durante 48 h para determinar la MS de campo parcial; posteriormente, se molieron en un molino de Willey (A. H. Thomas, Philadelphia, PA).
La MS total se determinó usando una estufa a 100°C durante 24 h con los procedimientos de la Asociación Oficial de Químicos Agrícola (AOAC, 2006). Finalmente, las muestras fueron incineradas en una mufla a 500°C para determinar la materia orgánica (MO) y el contenido de cenizas. Los contenidos de NDF y ADF se determinaron usando el método de Goering y Van Soest (1970). Las muestras también se analizaron para determinar el contenido de CP utilizando el método de Kjeldahl (AOAC, 2006). Los contenidos minerales se determinaron mediante espectrofotometría de absorción atómica (Galyean y May, 1996).
Cambios de peso y medidas corporales
Los PV y ECC de las vacas se midió y se registró. Los cambios diarios de PV se estimaron por diferencia entre dos PV semanales consecutivos. Las vacas se ordeñaron a las 08:00 y 20:00 h utilizando una ordeñadora en paralelo (Westfalia Surge, Bonen, DE). La leche producida se pesó y se registró con un lacto-metro Waikato MK V (Waikato, NZ). La PL diaria fue la suma de los dos ordeños del día. Las muestras de leche se tomaron en viales de 120 mL, que contenían una pastilla de Bromopol (Bromopol BSM2 D & F Control, San Ramón, CA) como conservador. Los contenidos de grasa, proteína, lactosa, sólidos totales se analizaron utilizando un MilkoScan FT120TM (Multispec, Foss Feed Technology Corp., Eden Prairie, CA).
La producción de proteína y grasa se estimó multiplicando el porcentaje de proteína y grasa, respectivamente, en la muestra por la PL diaria. La PL corregida al 4.0% de grasa de leche se estimó con la fórmula:
PLC= (0.4*PL) + [15* (PL* (Concentración de grasa /100)]
Análisis estadísticos
Los datos se analizaron con el procedimiento Mixed del Statistical Analysis System (SAS, 2012). Los PV preparto y los incrementos semanales de PV se utilizaron para estimar los cambios en el PV; mientras que el CMS, los cambios de PV, PL, y la eficiencia de PL fueron analizados con el procedimiento Mixed de SAS en un diseño completamente al azar con medidas repetidas en el tiempo. El modelo inicial incluyó los efectos fijos del tratamiento, semana, la interacción tratamiento x semana y vaca como término aleatorio. El modelo final, después de eliminar las covariables y las interacciones no significativas (p > 0.05) doble o triple, fue:
Yijkl = μ + TRATAMIENTOi + SEMANAj + TRATAMIENTO * SEMANAij + VACAk + Eijkl
Donde: Yijk es la variable de respuesta; μ es la media general; TRATAMIENTO es el efecto fijo del i-ésimo tratamiento (i = 1, 2); SEMANAj es el efecto fijo de la j-ésima semana (j = 1, 2, ..., 7); TRATAMIENTO * SEMANAij es el efecto fijo de la interacción del tratamiento x semana; VACAk es el efecto aleatorio de kth vaca; y Eijk es el término de error aleatorio.
Otros modelos similares, con diferentes interacciones y covariables, se utilizaron para analizar las concentraciones de CP, ADF, NDF y DM en las dietas. De los resultados obtenidos se reportan las medias de cuadrados mínimos y el error estándar de las medias. Cuando los efectos del TRATAMIENTO fueron significativos, se utilizó la opción PDIFF del procedimiento mixed para determinar las diferencias entre tratamientos. Para cada variable de respuesta se utilizó la mejor estructura de covarianza, de acuerdo con los criterios bayesianos de Akaike y Schwartz.
La estructura de covarianza más adecuada para el porcentaje de grasa y proteína, lactosa, y sólidos totales en leche fue la simetría compuesta; mientras que la estructura autorregresiva fue la mejor para CMS. La simetría de compuestos heterogéneos fue mejor para PL, eficiencia de alimentación, y rendimiento de grasa y proteína.
Resultados
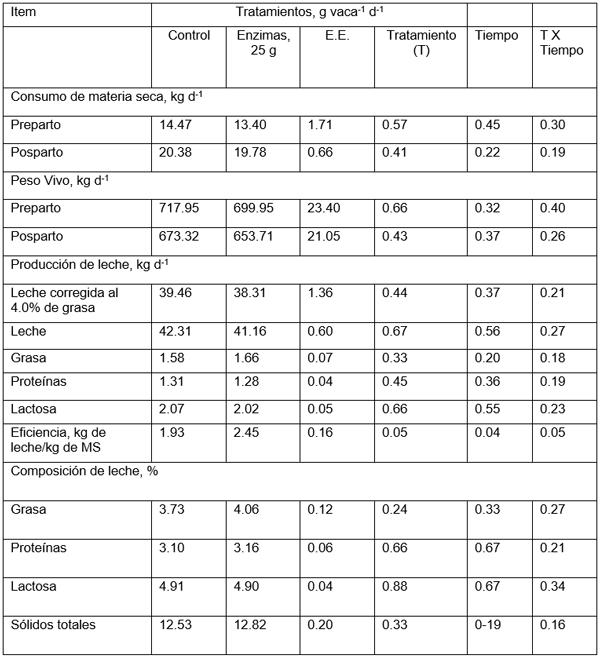
En el Cuadro 2 se presentan los consumos de materia en el periodo pre- y posparto, la producción de leche corregida al 4% de grasa, contenido de sólidos totales, grasa, proteína y lactosa de vacas Holsrein-Frieisian que consumieron dietas complementadas o no con enzimas fibrolíticas. La complementación dietética de 25.00 g animal-1 día-1 de enzimas fibrolíticas no afecto (p < 0.05) el consumo de materia seca de las vacas en el pre- y el posparto.
El Cuadro 2 muestra la producción de leche y la corregida al 4.0% de grasa de las vacas que fueron complementadas o no con 25.00 g animal-1 día-1 de enzimas fibrolíticas. La complementación dietética no influyó (p > 0.05) en la produccion de leche, ni tampoco en la corregida al 4.0% de grasa en las primeras siete semanas de lactancia.
El contenido de sólidos totales mostró un comportamiento similar a la producción de leche. La complementación con enzimas fibrolíticas no influyó (p > 0.05) en el contenido de sólidos totales de la leche (Cuadro 2). En la mismo Cuadro se observa que conforme transcurren las semanas después del parto, el contenido de sólidos en leche va disminuyendo. La disminución entre la semana uno y la siete es de aproximadamente 2.8% (Cuadro 2).
En el Cuadro 2 se muestra el contenido graso de la leche en porciento cuando las dietas fueron complementadas o no con 25.00 g de enzimas fibrolíticas durante siete semanas de lactancia. La complementación dietética no influyó (p > 0.05) en el contenido graso de la leche.
Los contenidos proteínicos de la leche (%) obtenido con la dieta complemente mezclada complementadas con 0.0 ó 25.0 g de enzimas fibrolíticas mostraron un comportamiento similar a la producción de leche. La complementación con enzimas fibrolíticas no influyó (p > 0.05) en el contenido porcentual de proteínas de la leche. También se observa que conforme transcurren las semanas después del parto, el contenido de proteínas de leche va disminuyendo. La dismunución entre la semana uno y la siete es de aproximadamente 26.00%.
El contenido porcentual de lactosa de la leche fue similar (p > 0.05) en ambos tratamientos (Cuadro2). En la mismo Cuadro se observa que conforme transcurren las semanas después del parto, el contenido porcentual incrementa. El incremento fue de aproximadamente 4.5% de la semana uno a la siete.
Discusión
Consumo de materia seca
El consumo de MS no fue afectado por la complementación con la enzima. Lo que quizás se deba a que la enzima mejoró la digestibilidad de MS, MO y los compuestos nitrogenados, incrementando la digestión de la fibra en el rumen. Sin embargo, los cambios no fueron suficientes para alterar la tasa de pasaje de las partículas ruminales (Zinn etal., 1999), lo que inhibió el incremento en el consumo de alimento. El incremento en el consumo de alimento de las vacas debido a la complementación con enzimas podría explicarse por el incremento en la tasa de pasaje de las partículas ruminales o por incremento en la digestión de la fibra, ninguno de los cuales se midieron en el presente experimento. Sin embargo, otros reportes de la literatura indicaron que los suplementos enzimáticos no aumentaron el CMS debido a que, aunque se incrementó la digestión de la fibra en el rumen (Feng et al., 1996), dicho aumento no fue suficiente para alterar la tasa de pasaje de las partículas ruminales (Zinn et al., 1999), lo que inhibió la expresión del consumo de alimento. Una de las bases de este postulado es el mejoramiento en la eficiencia alimenticia de las dietas, como se observa en el Cuadro 2.
La dieta se formuló con el 63.00% (alfalfa henificada y ensilado de Maíz) de forrajes, que posiblemente, mejoraron su digestibilidad o la digestibilidad de la fibra detergente neutro (FDN) con preferencia por el ensilado de Maíz (Cuadro 1).
Phakachoed et el. (2013) indicaron que las xilanasas incrementaron la digestibilidad de la fibra, y principalmente, de FDN y FDA in vitro del ensilado de Maíz. En las condiciones del estudio de Phakachoed et al. (2013) la dosis más apropiada fue de 2.0 µL g-1 de MS de ensilado de Maíz. En el presente estudio, la dieta preparto contenía aproximadamente 41.20% de ensilado de Maíz, que aparentemente, es un sustrato preferido por las xilanasas. Además, la dieta contenía 22.24% de heno de alfalfa, mayormente preferido por las celulasas, que tenía una digestibiildad de FDN de aproximadamente 33.0 %.
En el presente estudio, contrariamente, la adición del complejo enzimático no influyó en el CMS duante las primeras siete semanas de lactancia; lo que quizás se deba al contenido de únicamente 21.59% de ensilado de Maíz, que representó aproximadamente la mitad del mismo producto en la dieta preparto.
En adición, la dieta de lactancia se formuló con 43.78 % de alfalfa. Esto pudiera tener más afinidad con celulasas que con xilanasas. El análisis de laboratorio indicó que la alfalfa verde tenía aproximadamente 33.0% de FDN, con digestilidad superior al 60.0% lo que apoyó, posiblemente, el mantenimiento de las condiciones ruminales independientemente de la adición de enzimas a las dietas.
Holsthausen et al. (2011) indicaron que la adición de 0.5 y 1.0 mL kg-1 de MS de Fibrozyme no influyó en el CMS de vacas Holstein-Friesian en el inicio de la lactancia. Las dietas se formularon con ensilado de cebada, y heno y ensilado de alfalfa. El impacto de la enzimas no fue importante en el mejoramiento de la digestibilidad del ensilado y el heno de alfalfa. Por el contrario, fue más impactante en el ensilado de cebada, lo que se atribuyó al tipo de fibra. El CMS fue menor para la dosis de 1.0 y tendió a ser más bajo que 0.5 mL de Fibrozyme por animal-1 día-1 comparado con el Control. La diferencia se basó en que las enzimas fibrolíticas tienen el potencial de incrementar la digestibilidad de la fibra de los forrajes mejorando el CMS de la dieta.
En el presente estudio, el complejo enzimático utilizado fue una combinación de Aspergillus niger y Trichoderma viride con una actividad xilanásica de 100 UI g-1 y con potencial para liberar 1.0 µmol de xilosa g-1. Como se indicó anteriormente, se adicionaron 25.0 g animal-1 día-1 del complejo enzimático, que liberaron aproximadamene 25.0 µmol de xilosa que es equivalente a las dosis utilizadas en el estudio de Holsthausen et al. (2011).
Otros reportes de literatura indicaron resultados similares a los observados en el presente estudio. Romero et al. (2016) indicaron que la adición de xilanasa plus (1.0 mL kg-1 de dieta completa) no mejoró el CMS (23.5 vs. 22.6 kg d-1) de vacas en el inicio de la lactancia. Sin embargo, hubo a una tendencia a incrementar la PL corregida por grasa (39.46 vs 41.17 vaca-1 d-1) y la grasa de la leche (1.44 vs 1.48 %). Paralelamente, no se observaron efectos de la adición de enzimas en la digestibilidad de las dietas, producción de proteínas de la leche y lactosa, como ocurrió en el presente estudio, con una dosis similar de actividad xilanásica.
Contrariamente, otros estudios (Beauchemin et al., 2000; Dean et al., 2013) reportaron efectos positivos en el CMS con enzimas fibrolíticas endógenas con actividad alta de xilanasas. En el inicio de la lactancia, cuando las vacas se encuentran en balance energético negativo, el CMS es controlado por el llenado del tracto gastrointestinal (Dado y Allen, 1995); consecuentemente, las vacas deberían responder en menor grado al incremento en el CMS como una consecuencia en el mejoramiento de la digestibilidad de la fibra (Oba y Allen, 2000).
En un estudio publicado por Chow et al. (2008), se observó que las vacas alimentadas con ensilado de cebada con FDN de digestibilidad alta, no respondieron mejorando el CMS durante la parte media de la lactancia, sin embargo, las vacas tendieron a mejorar la ganancia de PV; lo que sugiere un mejoramiento del ambiente del tracto gastrointestinal. En el presente estudio, aunque no se determinó la digestibilidad de los forrajes en el tracto gastrointestinal, es posible inferir, que la diferencia a favor de la adición de enzimas, quizás se deba a un proceso parecido al observado por Chow et al. (2008).
Producción y calidad de la leche
El efecto de la complementación con enzimas fibrolíticas no influyó en la PL y tampoco en la PL corregida al 4.0% de grasa en las primeras siete semanas de lactancia (Cuadro 2). Esto pudo deberse a que, tanto la dieta Control como la adicionada con enzimas mostraron CMS similares durante dicho periodo. Como se discutió anteriomente, la falta de respuesta en el consumo de explicó por la afinidad de las celulasas por la alfalfa verde y las xilanasas por el ensilado de Maíz y que posiblemente, las últimas redujeron su actividad conforme disminuyó el nivel del ensilado de Maíz contenido en la dieta. El resultado final es que, teóricamente, ambas dietas proprcionaron la misma cantidad de nutrientes requeridos para síntesis de leche, como lo expresó el nivel similar de PL entre las dietas (Cuadro 2).
Los resultados obtenidos en el presente estudio coinciden con los observados por Refat et al. (2018), quiénes indicaron que la complementación con enzimas fibrolíticas derivada de Trichoderma reesei (FETR; mezcla de xilanasa y celulasa; AB Vista, Wiltshire, Reino Unido) aunque aumentaron linealmente la digestibilidad in vitro de la MS y tendieron al mejoramiento de la misma del ensilaje de cebada, no mostraron efectos en el CMS, la PL, producción de grasa de leche, leche corregida por grasa, y leche corregida por energía.
Resultados similares fueron observados por Holsthausen et al. (2011) y Chow et al. (2008) quiénes reportaron que no hubo efectos de la adición de enzimas con actividades xilanásicas en la PL. Lo anterior, sugiere que las diferecias en el mejoramiento del ambiente ruminal, si es que existieron no fueron suficientes para reflejar niveles de PL diferentes.
Otros reportes de literateratura demostraron efectos contradictorios a los del presente estudio. Eun and Beauchemin, (2005) reportaron incrementos en la producción de leche, proteína, grasa y lactosa de la leche con la adición de enzimas en dietas con alto y bajo contenido de heno de alfalfa y ensilado de cebada. Los incrementos se explicaron por un aumento en la degradabilidad de los forrajes en las dietas con contenidos bajos del mismo. Dicho mejoramiento promovió un CMS mayor a favor de la adición de enzimas con actvidad xilanásica, lo que suministro niveles deferentees de energía que fueron, posiblemente, utilizados en la fromación de leche.
La adición de productos comerciales con enzimas fibrolíticas exógenas a las vacas lecheras mejora la digestibilidad de los forrajes en el tracto total, principalmente, en dietas con contenidos bajos de forajes, particularmente, heno de alfalfa y ensilado de Maíz. También mejora la digestibilidad de la fibra de los otros componentes de la dieta. Lo anterior, sugiere un mejoramiento en la actividad ruminal de las enzimas fibrolíticas reforzando la idea de que las enzimas exógenas mejoran la actividad de la microflora del rumen, lo que resulta en un mejoramiento promedio del consumo de alimento.
En el presente estudio no se observaron respuestas en los rendimientos y contenidos (Cuadro 2) de sólidos totales grasa, proteínas y lactosa de la leche de vacas complementadas con 25 g animal-1 día-1 con enzimas fibrolíticas en el inicio de la lactancia. En concordancia con los resultados del presente estudio, varios reportes de la literatura han documentado que la complementación con enzimas fibrolíticas a vacas Holstein-Friesian no afectó la composición de la leche (Bernard et al., 2010; Arriola et al. 2011).
Por el contrario, Dyaa El-Din et al. (2013) observaron incrementos en el rendimiento de grasa, proteínas, lactosa y sólidos totales de la leche cuando las vacas se complementaron con 15 g animal-1 día-1 de Fibrozyme que es una dosis menor a la uilizada en el presente estudio. Los incrementos en el contenido graso lo explicaron por incrementos en la disponibilidad de energía y ácidos grasos para síntesis de grasa de la leche. La diferencia entre el estudio de Dyaa El-Din y el presente se basó en las dosis y los tiempos de suministro. En el presente estudio, se suministraron 25 g animal-1 día-1 mientras que en el estudio de Dyaa El-Din et al. (2013) se utilizaron 15 g animal-1 día-1 en un periodo de 12 semanas. La diferencia puede atribuirse a las condiciones ambientales diferentes entre los estudios.
El mejoramiento en la eficiencia lechera observada en el presente estudio puede atribuirse a un mejoramiento del ambiente ruminal propiciado por la enzima, que aunque no fue suficiente para demostrar la digestibilidad de los componentes de la dieta, mejoró el CMS. La reducción en el CMS (20.38 vs 19.78 kg) durante la lactancia explicó la eficiencia alimenticia en lugar de la PL o la ganancia de PV. Una tendencia similar fue observada por Holsthausen et al. (2011), quienes observaron que la desaparición ruminal de heno de alfalfa, MS, FDN, y FDA fueron mayores con 2.0 µL g-1 de forrajes en comparación con el grupo Control.
A pesar de que la adición del complejo enzimático no impactó positivamente el comportamiento de los animales durante la lactancia, el estudio sugiere que hay un mejoramiento en la efieciencia lechera de las vacas en condiciones d confinamiento.
Varias referencias del presente documento y los resultados del presente estudio, indican que los efectos de la adición de enzimas, como las usadas en el presente estudio, tienen resultados contradictorios a los indicados por las empresas expendedoras. Sin embargo, los reportes de las mismas empresas indican que los productos se venden considerablemente en los establos del país. Hasta donde los efectos potenciales de las enzimas pueden estar enmascarados por la dieta basal, el manejo del hato ó las condiciones ambientales locales, no lo sabemos. Por lo que consideramos, se seguirá trabajando con la visión de que las empresas tienen razón. Sin embargo, quedan preguntas de la efectividad de los productos en el mejoramiento del consumo de alimento y el suministro de nutrimentos para la formación de carne o leche. Una idea contraria, es que posiblemente, no haya motivos para incluir enzimas fibrolíticas exógenas en las dietas de vacas lecheras en el inicio de la lactancia.
Conclusión
Los datos indican que la adición de 25 g animal-1 día-1 de enzimas fibrolíticas con actividad xilanásica a la dieta no mejoran el consumo de alimento tres semanas antes del parto y siete semanas posparto. La producción y la composición de leche de vacas Hosltein-Friesian en las primeras siete semanas de lactancia no fueron impactadas por la adición de la enzima.
Cuadro 1. Ingredientes y composición química de las dietas experimentales ofrecidas durante el periodo preparto (21 días) y posparto (56 dí) adicionadas con 0.0 ó 25 g animal-1 día-1 con enzimas fibrolíticas a vacas Holstein-Friesian en confinamiento
Cuadro 2. Consumo de materia seca, producción y composición de la leche, y eficiencia lechera de dietas ofrecidas durante el periodo preparto (21 días) y posparto (56 días) adicionadas con 0.0 ó 25.0 g animal-1 día-1 de enzimas fibrolíticas a vacas Holstein-Friesian en condiciones de confinamiento