Granos de Destileria y Sindrome de Depresion Grasa en la Leche
Publicado: 9 de octubre de 2012
Por: Fernando Díaz-Royón | DVM Álvaro García | Dairy Specialist, DVM, Ph.D.
Introducción
La grasa es el componente de la leche más fácilmente manipulable con la dieta, pudiendo llegar a variar en un rango de 3 unidades porcentuales (Jenkins y Lock, 2008). La nutrición representa el factor ambiental más importante que afecta a la grasa de la leche y es una herramienta para alterar su composición en ácidos grasos. La depresión de la grasa láctea ocurre cuando dietas específicas provocan una disminución en el contenido y rendimiento de grasa. Desde un punto de vista económico, la formulación de raciones para vacuno lechero a menudo indica incluir en las dietas tantos granos de destilería (GD) como sea posible. Para poder conseguir un balance adecuado de nutrientes, el nivel máximo de inclusión recomendado ha sido de un 20% de la materia seca (MS) de la dieta (Schingoethe, 2009). En la práctica muchos productores y técnicos piensan que la alta concentración en lípidos de los GD provoca caída de la grasa láctea (Diaz-Royon y col., 2011). Debido al alto contenido en fibra de los GD, los técnicos pueden verse tentados en usarlos para remplazar parte de la fibra neutro detergente (FND) del forraje. Sin embargo, como consecuencia de su pequeño tamaño de partícula, la FND de los GD no es "efectiva", es decir, tiene poca capacidad de estimular la rumia y mantener valores normales de grasa láctea (Schingoethe y col., 2009). Estos factores pueden originar la aparición de acidosis ruminal, la cual puede agravar los efectos negativos que la alta carga de ácidos grasos (AG) poliinsaturados ocasiona sobre la caída de la grasa láctea. Estos efectos han sido observados por Ciriac y col. (2005) en South Dakota State University. Estos autores encontraron una disminución lineal en la concentración de grasa láctea (3.34 a 2.85%) sin disminución de la producción de leche, cuando los granos secos de destilería con solubles (DDGS) sustituyeron 7, 14 y 21% del ensilado de maíz de la dieta, aun manteniendo constante la FND de las dietas (32%).
Fibra efectiva
El contenido del rumen es mezclado durante la rumia, siendo las partículas largas y menos digestibles del forraje regurgitadas, masticadas y vueltas a tragar. Este ciclo se repite cada vez que la vaca ingiere partículas largas de forraje. Durante este proceso, la cutícula de protección que cubre los fragmentos de las plantas es macerada, y así los microrganismos ruminales pueden acceder al contenido celular altamente digestible. Las partículas de alimento terminan este ciclo cuando adquieren el tamaño y densidad necesarios para sedimentar a la parte inferior del rumen y/o pasar al intestino. Además de disminuir el tamaño de las partículas, la rumia estimula la producción de saliva. Debido a su alto contenido en bicarbonato, la saliva es esencial para neutralizar la acidez producida por la fermentación microbiana en el rumen. Los carbohidratos estructurales (ej., FND) inician la rumia a través de estímulos físicos. Esos carbohidratos, forman una masa flotante fibrosa en el rumen que juega un papel muy importante para la salud y funcionalidad de este órgano. Esta masa frota las paredes ruminales, este estímulo físico es necesario para que el rumen se contraiga y comience el ciclo de la rumia. Cuando las partículas ingeridas son muy pequeñas se compromete la formación de esta masa. En su ausencia, la motilidad ruminal se compromete, la retención de la ingesta disminuye y al mismo tiempo, la digestibilidad de la dieta. Esta masa de partículas podría compararse a un "filtro de forraje" que enlentece las partículas lo suficiente para que los microrganismos del rumen las degraden. La disminución de las contracciones del rumen se acompaña de reducción en la masticación del alimento y en consecuencia, menor producción de saliva. Debido a esto se produce una disminución de la capacidad de neutralizar la acidez ruminal. Esta es la razón por la cual la concentración de FND de la dieta está poco relacionada con la "efectividad" de esta fibra. Lo que importa realmente, es el tamaño de partícula de la misma. Trabajos de investigación demostraron que con pH ruminal por encima de 6, el porcentaje de grasa láctea en vacas Holstein fue 3.5 o superior (Grant et al., 1990). Un pH ruminal relativamente bajo pero sin que llegue a causar acidosis clínica, puede ocasionar cambios en las rutas de bio-hidrogenación ruminal (BH). Estos cambios provoca un aumento en la formación de ácido linoleico conjugado (CLA) trans-10, cis-12 y otros intermediarios de la BH que se han identificado como inhibidores de la síntesis de grasa en la glándula mamaria (Lock y col., 2006).
Fermentación ruminal
La digestión ruminal de la grasa incluye sucesivamente la hidrólisis e hidrogenación de los AG insaturados. Por ejemplo, la BH del ácido linoleico (cis-9, cis-12 18:2) tiene lugar en tres reacciones: en un primer paso se produce una isomerización que da lugar a la síntesis de cis-9, trans-11 CLA, y posteriormente dos reducciones sucesivas, primero a ácido oleico (trans-11 C18:1) y después a ácido esteárico (C18:0; Harfoot y Hazelwood, 1988). Pero según la "Teoría de la Bio-hidrogenación" propuesta por Bauman y Griinari en 2001, bajo ciertas condiciones, el ambiente ruminal está alterado y parte de la BH se produce mediante una ruta que genera intermediarios específicos, como trans-10, cis-12 CLA y trans-10 18:1 CLA (Figura 1).
El paso al intestino de estos intermediarios depende de la tasa de pasaje ruminal, la capacidad de BH bacteriana, y la concentración y perfil de los AG poli-insaturados presentes en el rumen (Lock y col., 2006). Una vez que han sido absorbidos en el intestino, mediante la circulación alcanzan la glándula mamaria, donde ocasionan una disminución en la expresión de enzimas lipogénicas, y por lo tanto un descenso en la síntesis de grasa láctea. Los trastornos ruminales son provocados por la actividad antibacteriana de los AG insaturados, los cuales penetran dentro de la membrana de algunas especies bacterianas y ocasionan la desorganización de los fosfolípidos, con el consiguiente daño celular (Jenkins, 2002). Debido a que los AG específicos que causan disminución de la producción de grasa son producidos durante BH de los AG poli-insaturados, es lógico pensar que la cantidad inicial de linoleico y quizás linolénico podría estar relacionada con la cantidad de intermediarios producidos (Jenkins y Lock, 2008). En conclusión, para producir depresión de la grasa láctea, son necesarias dos condiciones; una de ellas es la presencia de AG insaturados en el rumen, la otra, un ambiente ruminal alterado que ocasione una BH incompleta de estos.
La población microbiana del rumen es capaz de degradar y fermentar carbohidratos y proteínas para producir ácidos grasos volátiles (AGV). Al elevar la concentración de AG en el rumen se provocan cambios en la distribución de la población microbiana. Estos cambios pueden afectar la fermentación de los carbohidratos causando un descenso en la proporción acetato: propionato, y una posible reducción en la digestión de la fibra (Jenkins, 2002). En la tabla 1 se presenta un resumen de los resultados obtenidos en 9 experimentos que compararon raciones con diferentes niveles de inclusión de GD. La concentración relativa (moles/100 moles) de AGV lipogénicos (acético y butírico) no sufrió variaciones significativas en 5 de los 7 experimentos evaluados. Sin embargo, en los estudios de Hippen y col. (2010) y Ranathunga y col. (2010) se produjo una disminución en la concentración relativa de acetato en el rumen. En la prueba de Hippen y col. (2010), además apareció una reducción en la producción total de AGV con las dietas que contenían 20% de DDGS.
Según los autores, las principales causas serían la baja cantidad de fibra efectiva y la presencia de AG insaturados. Ranathunga y col. (2010) sugirieron que el descenso lineal en la concentración relativa de acetato observada al incrementar los DDGS en las dietas, podría atribuirse a una disminución en el consumo de FND procedente del forraje o a mayores concentraciones de AG insaturados en estas dietas. Esta menor concentración de acetato, también provocó una disminución lineal en la proporción acetato: propionato. Ambos estudios encontraron una alteración en el ambiente ruminal, pero sólo el experimento de Hippen y col. (2010) mostró un ejemplo típico de depresión de la grasa láctea, con caída de la concentración y rendimiento de grasa.
Dietas experimentales
De acuerdo con el NRC (2001) los GD contienen un 10% de grasa sobre materia seca (MS), sin embargo, los análisis publicados por Dairy One Lab (4819 muestras, 2011), muestran un contenido medio de 12.6% (9.4 a 15.7%). Estos datos reflejan una alta variabilidad en su contenido graso. El AG linoleico es el más abundante, representando el 53.7% de los AG totales (Moreau y col., 2011). El gráfico 1 refleja el porcentaje de incremento del AG linoleico en las dietas al aumentar la inclusión de GD. Los datos proceden de 4 experimentos, en los que el ensilado de maíz representó el forraje más abundante de la ración. La cantidad total de AG linoleico en dietas sin GD fue de 294 g (Singoethe, 1999) a 439 g (Leonardi y col., 2005). La dieta con mayor contenido en linoleico fue la de Leonardi y col. (2005) con 595 g, a un nivel de inclusión de GD del 15% (MS). Las vacas que consumen dietas compuestas por ensilado de maíz como fuente forrajera única o predominante, parecían ser más susceptibles a causar caída de la grasa, particularmente cuando además eran suplementadas con grasas insaturadas (Staples, 2006). Ruppert y col. (2003) mostraron que en dietas basadas predominantemente en ensilado de maíz, al suplementar con grasa se producía una caída de la grasa de la leche. Sin embargo, esta situación fue corregida al sustituir parcialmente el ensilado de maíz por ensilado de alfalfa.
De los 9 experimentos analizados en la tabla 1, sólo en el estudio de Hippen y col (2010), en el cual compararon una dieta control con otra que contenía un 20% de DDGS, se constató una disminución en la producción de leche. En dos de los experimentos, al incluir GD aumentó la producción de leche. Leonardi y col. (2005) obtuvieron un incremento lineal en la producción láctea al aumentar la inclusión de DDGS. Anderson y col. (2006) también obtuvieron mayor producción de leche en dietas que incluían un 10 o un 20% de GD que con la dieta control. Además, Anderson y col. (2006) no encontraron diferencias en producción láctea entre las dietas que contenían GD húmedos o secos, o al variar su nivel de inclusión (10 o 20%). Estos resultados coinciden con los publicados en dos estudios anteriores (Hippen y col., 2003; Kalscheur y col., 2004) en los que generalmente los GD produjeron una disminución en la producción láctea exclusivamente cuando se incluyeron en las dietas a niveles superiores al 20%. El aumento de la producción de leche al incluir GD obtenido en los estudios de Leonardi y Anderson puede ser debido a que las dietas no contenían la misma cantidad de grasa, ya que las raciones con GD contenían más grasa que las raciones control. Sin embargo, la dieta control de Hippen y col. (2010) incluyó una fuente de grasa inerte con el objetivo de igualar el contenido graso de la dieta con DDGS. En los 6 estudios restantes no se encontraron diferencias en producción láctea estadísticamente significativas.
Cambios en la grasa láctea
La grasa láctea esta compuesta por una compleja mezcla de lípidos, aunque un 97% de estos son triglicéridos (Christie, 1995). Estos lípidos son sintetizados a partir de "precursores" procedentes de la circulación periférica (60%) o sintetizados "de novo" en la glándula mamaria (40%; Chilliard y col. 2000). Los AG con longitud de cadena de 4 a 14 carbonos (C), y una porción de los de 16 C son derivados de síntesis de novo en la glándula mamaria a partir de acetato, y en menor proporción de betahidroxi- butirato. El resto de los AG de 16 C, y todos los de 18 C y de cadenas más largas son derivados de AG circulantes, procedentes de la absorción de lípidos de la dieta o de la movilización de grasa corporal (Bauman y Griinari, 2001). Durante el síndrome de depresión láctea, se produce un descenso en la secreción de la mayoría de los AG, pero además se modifica la composición de la grasa, debido a que esta reducción afecta principalmente a AG sintetizados de novo (David y Brown, 1970). Como resultado, en la leche disminuye la concentración de AG de cadena corta y media e incrementa la concentración de AG de cadena larga (Bauman and Griinari, 2001).
El incremento en la grasa láctea de intermediarios de la BH como el trans-10, cis-12 CLA es indicativo de trastornos ruminales que se han producido en las rutas de BH de AG poli-insaturados. En vacas alimentadas con una dieta baja en fibra y suplementada con aceite de girasol, Griinari y col. (1999) observaron una relación estrecha entre el porcentaje de grasa en leche y su contenido en trans-10, cis-12 CLA. Además, los cambios que se produjeron en la composición de AG de la leche durante la infusión abomasal de trans-10, cis-12 CLA fueron consistentes con los observados en dietas que produjeron caída de grasa (Bauman y Griinari, 2001). Tan solo en 3 de 8 experimentos de la tabla 1, se produjo un aumento de este isómero en la leche en dietas que contenían GD. Excepto en el experimento de Hippen y col. (2010), en el que el aumento del isómero esta relacionado con caída de la grasa, en los otros dos estudios (Abdelqader y col., 2009; Anderson y col., 2006) este incremento en la concentración de trans-10, cis-12 CLA no fue acompañado de una reducción en la producción de grasa. A pesar de que ambos estudios reportan diferencias estadísticamente significativas en la concentración de este isómero en la leche, las bajas concentraciones a las que aparece pueden ser insuficientes para provocar una disminución en la síntesis de grasa.
Ocasionalmente las dietas provocan un incremento en la producción láctea sin cambios en el rendimiento graso, y como consecuencia de esto el porcentaje de grasa en la leche disminuye. Esto no representa síndrome de depresión grasa, ya que la síntesis de la misma no ha sido afectada. En un tercio de los experimentos de la tabla 1 se redujo el porcentaje de grasa en la leche. Hippen y col. (2010) encontraron una disminución tanto del porcentaje como del rendimiento graso. Por el contrario en los experimentos de Abdelqader y col. (2009), y Leonardi y col. (2005) se observó una disminución del porcentaje sin diferencias en la producción total de grasa. Los resultados obtenidos por Hippen y col. (2010) coinciden con los publicados en un meta análisis de Kalscheur (2005), donde se observó que los GD provocaban caída de la grasa exclusivamente cuando las dietas contenían menos del 50% de forraje o del 22% de FNDfo. La dieta con DDGS de Hippen y col. (2010) contenía un 45% de forraje y un 19.3% de FNDfo. La cantidad de ración total mezclada (RTM) recuperada en las dos bandejas superiores del Separador de Partículas Penn State (SPPS, 4 bandejas, 3 cribas) fue similar, pero la dieta que contenía DDGS tuvo aproximadamente un 3% menos de material retenido en la tercera bandeja y un 3.8% más en la bandeja inferior. A pesar de que todas las fracciones se encuentran dentro de las recomendaciones para RTM de vacas de alta producción (Zebeli y col. 2007), el paso de material de la tercera a la cuarta bandeja representa un mayor contenido en alimentos más rápidamente fermentables o con una mayor tasa de pasaje. Esto ha sido asociado con una disminución en la concentración y rendimiento de grasa de la leche (Grant y col., 1990; Beauchemin y col., 1997; Kononoff y col., 2003; Soita y col., 2005). Estos resultados son similares a los obtenidos por Nydam y col. (2008) en un estudio epidemiológico en el que estudiaron los factores de riesgo que contribuyen al síndrome de depresión grasa en granjas comerciales que utilizaban monensina. La separación de muestras de RTM mediante el SPPS (3 bandejas, 2 cribas) mostró una relación significativa entre el porcentaje de grasa y el tamaño de partícula. Esta relación fue positiva para la criba del medio y negativa para la bandeja inferior. Sin embargo, el porcentaje de grasa no estuvo relacionado con la fracción retenida en la criba superior. En el experimento de Anderson y col. (2006) se reportó un aumento en la producción de grasa acompañado además de un aumento en el rendimiento de leche. Los otros 6 experimentos evaluados no revelaron efectos significativos en el rendimiento o porcentaje de grasa al incluir GD en las dietas.
La adición de GD en las dietas disminuyó las proporciones relativas (g/100 g de AG) de AG de cadena media e incrementó la concentración de AG de cadena larga en leche en 4 de los 8 estudios analizados en la tabla 1. Además, en dos de ellos, también se produjo una disminución en los AG de cadena corta. Estos cambios pudieron deberse a tres razones:
1. Reducción de la síntesis de novo de AG debido al efecto de intermediarios de la BH sobre enzimas lipogénicas de la glándula mamaria (Bauman y Griinari, 2001)
2. Mayor secreción de AG de cadena larga procedentes de precursores sanguíneos, provocando un efecto de dilución (Chilliard y col. 2000).
3. Disminución de la disponibilidad de acetato y beta-hidroxi-butirato para la síntesis de AG de la glándula mamaria (David y Brown, 1970).
Conclusión e implicaciones
Algunas recomendaciones prácticas pueden extraerse de estos resultados con el objetivo de maximizar la inclusión de GD en las dietas sin producir efectos negativos en la producción o concentración de la grasa láctea.
1. Aunque la formulación de dietas con concentración adecuada de FND es importante, debe ir acompañada con una evaluación del tamaño de partícula de la ración total mezclada.
2. Añadir buffers a la dieta (ej. bicarbonato) puede disminuir los efectos que produce un pH ruminal bajo, pero su efecto es transitorio y no debe sustituir a un tamaño de partícula adecuado de la ración.
3. La cantidad y composición del resto de los alimentos incluidos en la dieta indica la cantidad de GD que se pueden añadir a esta.
4. Incluso con tamaño de partícula adecuado, la inclusión de más del 10 % de GD húmedos puede ser arriesgado cuando la ración esta compuesta por un 50% o más de otros productos fermentados que proceden del maíz, como son el ensilado de maíz o el maíz alta humedad.
Figuras, graficos y tablas
Figura 1. Bio-hidrogenación ruminal del ácido linoleico.
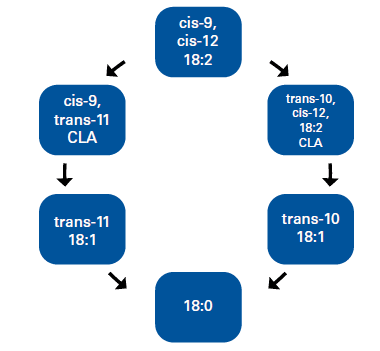
Nota: Adaptado de Griinari y Bauman (1999).
Figure 2. Incremento porcentual de AG linoleico al aumentar la inclusión de GD en las dietas.
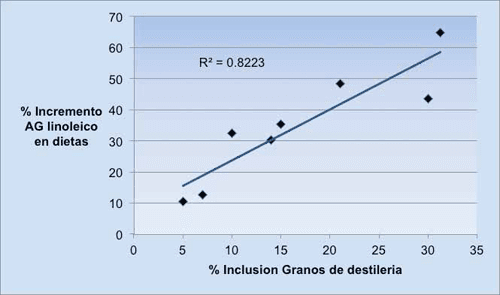
Fuente: Abdelqader y col. (2009); Leonardi y col. (2005); Ranathunga y col. (2010) y Schingoethe y col. (1999).
Tabla 1. Evaluación de la respuesta productiva en dietas con diferentes niveles de inclusión de granos de destilería.
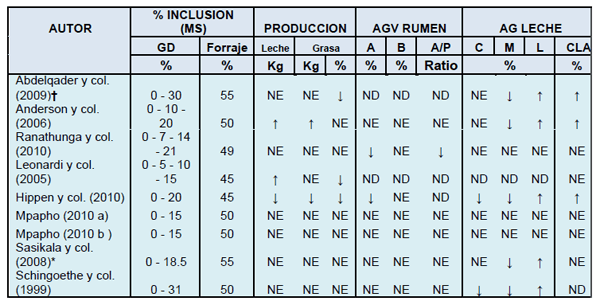
Abreviaturas: MS (Materia Seca); AGV (Acidos Grasos Volátiles); AG (Acidos Grasos); GD (Granos de destilería); A (Acetato); B (Butirato); P (Propionato); C (cadena corta); M (cadena media); L (cadena larga); CLA (Acido linoleico conjugado); NE (efecto no significativo, P>0.05); ND (datos no disponibles), ↑ (incremento); ↓ (disminución). †: La dieta control contiene un 16.3% (MS) de GD altos en proteína (3.3%grasa sobre MS); *: En este estudio se consideró dieta control aquella que contenía 10% de condensados de destilería.
Temas relacionados:
Autores:

South Dakota State University

GPS Dairy Consulting
Recomendar
Comentar
Compartir

Royal Agrifirm Group
27 de noviembre de 2012
Fernando y Alvaro, muy buen artículo. Considero muy importante que los técnicos no estigmaticemos insumos muy útiles para la nutrción porque utilizados correctamente (basados en información de calidad) dan muy buenos resultados.
Recomendar
Responder

GPS Dairy Consulting
16 de octubre de 2012
El separador de partículas de Penn State permite evaluar el tamaño de las partículas en la racion completa mezclada (RCM).El material retenido en cada criba debe seguir esta distribución:
• Criba superior (19mm): 2 - 8%
• Criba media (8mm): 20 - 30%
• Criba inferior (1.8mm): 20 - 30%
• Bandeja baja: = 20
Además, el separador de partículas también permite comprobar si los animales están seleccionando las partículas finas en la RCM. Para ello es necesario evaluar la uniformidad de la RCM a lo largo del tiempo (cada 6 horas) y en los rechazos. El material retenido en cada criba del separador de partículas no debe variar más de 5 unidades porcentuales con respecto a la RCM recién distribuida.
Recomendar
Responder

14 de octubre de 2012
Ha sido un interesante experiencia. La Saliva es fundamental para un proceso normal de rumia en el vacuno. Sugiero que podría considerarse condiciones ajenas ambientales como el stress calorico en vacas, que tambien originan deshidrataciòn y menor producción de saliva en vacunos de leche. le comento que esto sucede en estos momentos en zonas áridas como Majes Siguas, Peru, a 1430 msnm. costa perteneciente a la franja norte del desierto de Atacama. El ganado vacuno lechero Hosltein al pastoreo pasa alrededor de 7 horas en pleno pastoreo de alfalfa, con una radiaciòn solar de IUV : 8 y temperatura que oscila entre 29 a 33 º C, sin sombras y agua fresca.. Se ve defecaciones con burbujas producto de acidosis ruminal por falta de saliva..El ganado vacuno jadea demasiado para refrescarse baja estas condiciones, sigo investigando esto...Felicitaciones.
Recomendar
Responder


21 de noviembre de 2012
Para el caso del Afrecho de Cerveceria, animales que estan consumiedo 15Kg de Materia Humeda (MS 40% aprox), cual debe ser la inclucion de Carbonato de Calcio, para evitar problemas de PH en el Rumen?
Recomendar
Responder
16 de octubre de 2012
cual seria el tamaño ideal de particula para bovinos. exelente documento
Recomendar
Responder

13 de octubre de 2012
Muy buen articulo. Gracias por mantenernos actualizados.
Recomendar
Responder
12 de octubre de 2012
Hola Amigos:
LOs felicito por compartir esta excelente información, los invito a que sigamos adelante, de mi parte muchisimas gracias porque me ha sido de gran utilidad, por estar inmerso en estos menesteres pecuarios, felicidades.-
Recomendar
Responder
11 de octubre de 2012
La información esta muy bien documentada...quienes nos ocupamos del tema alimentario animal, nos permite consultar estas investigaciones... Gracias por compartirlas..
Recomendar
Responder
10 de octubre de 2012
Felicidades por su importante articulo, muchas gracias por compartir esta informacion de mucha importancia.
Recomendar
Responder

Danone
10 de octubre de 2012
Muy buena revisión, muchas gracias
Jaime Alcañiz
Recomendar
Responder

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar
