Avances científicos recientes (2023, Q1) acerca del uso del nitrógeno, proteína y aminoácidos en vacuno lechero
Publicado: 24 de julio de 2023
Por: Alfredo J. Escribano. ORFFA.
Resumen
El presente trabajo tiene por objetivo la extensión de conocimiento en este campo y la complementación del artículo previamente publicado por BM editores y el mismo autor, titulado “Nitrógeno en vacuno lechero”. Para tal fin, se han compilado y resumido algunas de las novedades científicas que nos ha traído el año 2023 hasta la fecha, organizados en bloques de mensajes clave (take home messages) que el autor considera de relevancia para el sector.
Figura 1. Resumen visual del trabajo
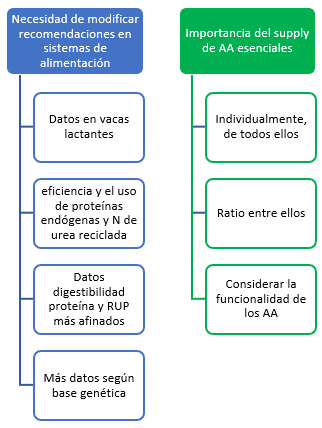
1. NECESIDAD DE MODIFICAR LAS RECOMENDACIONES EN LOS SISTEMAS DE ALIMENTACIÓN: de extrapolaciones y valores fijos a recomendaciones más basadas en datos abundantes y en vacas lactantes. Fuente: Extracto traducido de Silva & Oliveira (2023).
Los sistemas de alimentación (recomendaciones) actuales presentan limitaciones, y por tanto, sesgo, lo que resulta de gran interés desde el punto de vista económico y medioambiental. En particular, y parafraseando a Silva & Oliveira (2023), en el pasado se ha asumido que el requerimiento neto de proteína para el mantenimiento (NPM) es la suma de la excreción fecal y urinaria de proteína endógena (EPFU) y las pérdidas de proteína por descamación (Owens, 1987; Lapierre et al., 2020). Un enfoque alternativo es estimar la EPFU mediante la extrapolación de la intercepción-regresión entre la ingesta de N y la excreción urinaria y fecal total de N (NFU; Swanson, 1977; Owens, 1987). El propio NRC (2001) adoptó el EPFU de Swanson, lo cual, puede conllevar una subestimación de NPM ya que se llevó a cabo una extrapolación de datos en vacas no lactantes, a vacas en lactación, cuya ingesta y excreción de N por masa corporal es mayor.
El nuevo modelo NASEM (2021) parece haber mejorado la predicción del rendimiento de proteína de la leche en comparación con NRC (2001) al adoptar los supuestos de Lapierre et al. (2020) para predecir EPFU, pero los datos de Lapierre et al. en vacas lecheras son escasos, y NASEM no ha testado tales recomendaciones con valores observados.
En cuanto al requerimiento de proteína para lactación y de la eficiencia del uso de la misma, los diferentes sistemas de alimentación varían en cuanto a su enfoque, convergiendo la mayoría de ellos en valores en torno a 0,67-0,70.
Bajo este contexto de imprecisión, llamémoslo así, o de necesidad de afinar, Silva & Oliveira (2023) publicaron recientemente su trabajo titulado “Un nuevo sistema de requerimiento de proteína para vacas lecheras” en el que desarrollaron un nuevo sistema factorial para contabilizar la eficiencia de NPM y MP para lactación a partir de un metaanálisis de pruebas de balance de N de vacas lecheras lactantes. Estimaron la EPFU (NPM) a partir de la intersección de una ecuación no lineal entre la ingesta de N y las excreciones urinarias y fecales combinadas de N. El modelo propuesto proporcionó una estimación robusta de EPFU. Propusieron un EMPL fijo calculado como una proporción del N metabolizable (ingesta de N menos N fecal y urinario) que se secretaba en la leche. Dicho sistema predijo el requerimiento de MP para lactancia y mantenimiento con mayor precisión que varios modelos de nutrición de ganado lechero.
Qué proponen estos autores:
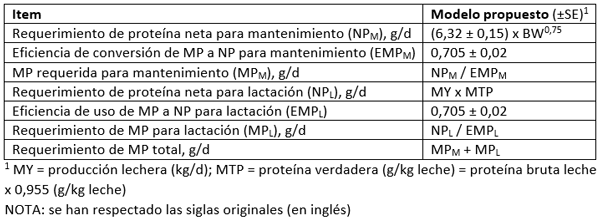
Estos autores definen el NPM como la suma de la excreción fecal y urinaria de proteína endógena, y la estiman a partir de la intersección entre la ingesta de N y la excreción fecal y urinaria de N total de forma combinada (6,32 ± 0,15 g de proteína/kg de peso corporal metabólico; n = 807 medias de tratamiento; r = 0,91).
Por su parte, calcularon la EMPL como una proporción (ingesta de N, menos el N excretado en las heces y la orina) de la excreción en leche (EMPL fijo de 0,705 ± 0,020), predeciendo el requisito de PM para la lactancia y el mantenimiento con mayor precisión que varios modelos de nutrición de ganado lechero de América del Norte y Europa, incluidos INRA (2018) y NASEM (2021). Solo los modelos NRC (2001), CNCPS 6.5 y Feed into Milk (2004) tuvieron una precisión similar para predecir el requisito de PM.
Tabla 1. Traducción de tabla 4 de Silva y Oliveira (2023). Descripción del sistema propuesto para proteína neta (NP) y requerimientos de MP para mantenimiento y lactación en vacas de leche
Qué falta aún; pasos hacia delante: los investigadores reconocen que todavía se necesitan modelos predictivos más refinados de digestibilidad intestinal para la proteína no degradable del rumen y la proteína microbiana para reducir los sesgos de evaluación para predecir los requisitos de MP de las vacas lecheras. Es preciso considerar aspectos que pueden afectar la estimación de los requisitos de mantenimiento, como:
- la eficiencia y el uso de proteínas endógenas y N de urea reciclada por parte de los microbios del rumen (Lapierre et al., 2016)
- tener en cuenta más factores involucrados en las pérdidas fecales endógenas de N, como el contenido de NDF y la tasa de fermentación de los carbohidratos de la dieta (Marini et al., 2008)
- generar más datos con las diferentes bases genotípicas: datos insuficientes de B. taurus × B. indicus (n = 26 medias de tratamiento) para vs. B. taurus (principalmente Holstein)
- afinar en las digestibilidades intestinales para proteína microbiana y RUP, ya que los rangos son amplios: digestibilidad intestinal de la proteína microbiana varía del 57 al 87 % según la proporción de bacterias/protozoos/hongos y el método adoptado (Jouany, 1996; Larsen et al., 2001; Fonseca et al., 2014; Fessenden et al., 2017), mientras que la digestibilidad intestinal total de AA RUP ha oscilado entre 52 y 94 % según el alimento, el procesamiento del alimento y el método (es decir, incubación de residuos en enzimas digestivas in vitro o en bolsas móviles insertadas en el duodeno) (White et al. , 2017).
2. EL SUPPLY DE AMIMOACIDOS ESENCIALES Y LAS RACIONES REDUCIDAS EN PROTEÍNA
2.1 Balanceo de raciones para nitrogeno ruminal y aminoácidos esenciales (EAA) (fuente: Balancing dairy cattle diets for rumen nitrogen and methionine or all essential amino acids relative to metabolizable energy; R. J. Higgs et al., 2023)
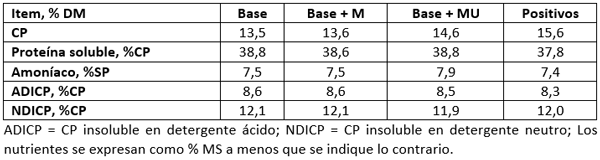
El objetivo de este estudio fue evaluar la sensibilidad de una nueva versión del Sistema Neto de Carbohidratos y Proteínas de Cornell (CNCPS) para formular dietas para N ruminal, Met y todos los AA esenciales (EAA). Sesenta y cuatro vacas lecheras de alta producción fueron asignadas aleatoriamente a 1 de las 4 siguientes dietas en un estudio longitudinal de 14 semanas:
- proteína metabolizable (MP), Met y N ruminal (Base) limitados
- Met adecuado pero MP y N ruminal limitados (Base + M)
- Met y N ruminal adecuados, pero MP limitado (Base + MU)
- MP, N ruminal adecuados y balanceados para todos los EAA (Positivos)
Tabla 2. Traducción de tabla 1 de Higgs et al. (2023) “Ingredientes y composición química de las dietas”
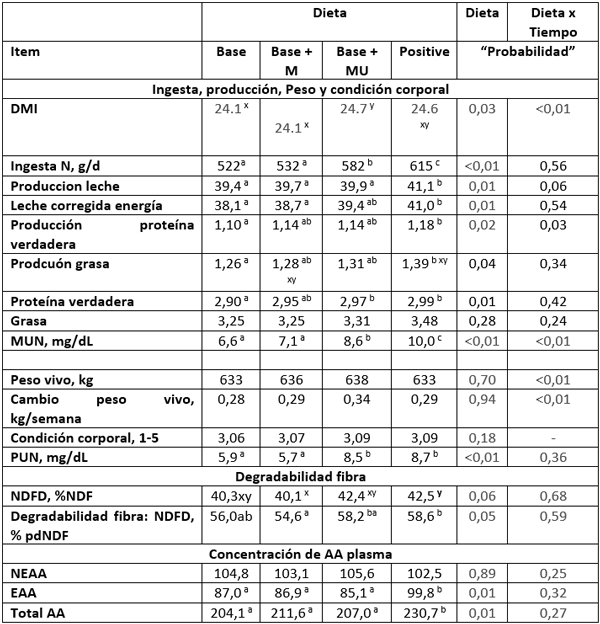
El estudio demuestra cómo se observan mejoras en los rendimientos de leche y componentes de la leche, ECM y degradación de la fibra cuando las dietas se formulan y suministran para cumplir con los requisitos de N ruminal y se mejoran aún más cuando se equilibra el suministro de EAA en función de la EM. Además, este estudio demostró que la aplicación de la versión actualizada del CNCPS es sensible en la predicción de cambios en el suministro de N, MP y EAA en el rumen, proporcionando una forma para que los usuarios mejoren el equilibrio de EAA en sus dietas y reduzcan los niveles excesivos de N en comparación con versiones anteriores del modelo.
Tabla 3. Integración y traducción de principales resultados de Higgs et al. (2023)
2.2. Debemos cuidar el suministro de todos los EAAs
2.2.1. Efectos de los desequilibrios entre EAA, escasez de suministro y consecuencia sobre el catabolismo de los mismos (y sobre la eficiencia de uso del N). Resumen de Van den Bossche et al. (2023). Effect of supplementing rumen-protected methionine, lysine, and histidine to low-protein diets on the performance and nitrogen balance of dairy cows. Journal of Dairy Science, Volume 106, Issue 3, 1790-1802
Hablando de aminoacidos concretos, recientemente, Van den Bossche et al (2023), evaluaron el efecto de suplemento de metionina, lisina e histidina protegidas en el rumen a las dietas bajas en proteínas sobre el rendimiento y el balance de nitrógeno de las vacas lecheras. Para examinar esto, se llevó a cabo un ensayo bajo las siguientes condiciones:
- período de adaptación pre-experimental de 3 semanas (dieta 16,5% de proteína cruda), seguido de
- un período de depleción de 4 semanas, en las que 39 vacas (promedio ± desviación estándar: 116 ± 29,3 d en leche, 1,8 ± 1,2 lactanciones, 638 ± 73,2 kg de peso corporal, y 32,7 ± 5,75 kg MY/d) recibieron una dieta baja en proteínas (CTRL) (14,5% de proteína cruda)
- asignación de animales de forma aletaoria en 3 grupos experiementales durante 7 semanas teniendo en cuanta paridad, concentración plasmática de Histidina y proddución láctea:
- AA período de 7 semanas; (1) CTRL
- (2) CTRL + RP-Met + RP-Lys (MetLys)
- (3) CTRL + RP-Met + RP-Lys + RP-His (MetLysHis)
- Dosificación de los aminoacidos protegidos en base a los necesidades de AA digestible: 2,4%, 7,0% y 2,4% para Met, Lys e His, respectivamente
Tabla 4. Resultados en periodo con suministro de AA protegidos. Extracto de Van den Bossche et al. (2023) traducido
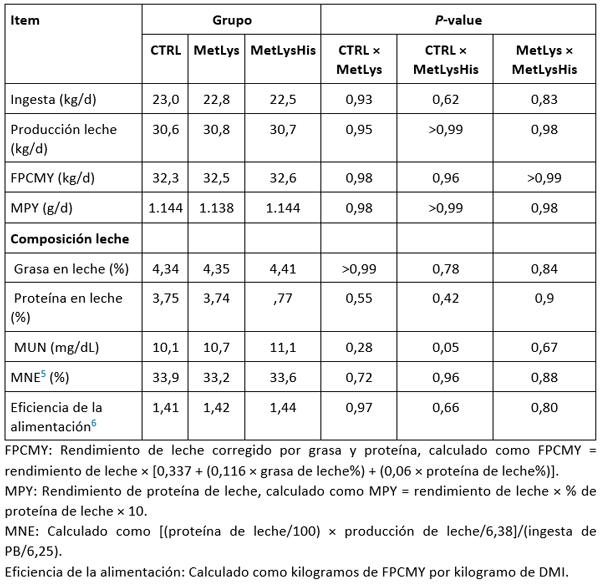
Tabla 5. Resultados medido durante la última semana de cada período (depleción, RP-AA, y cross-back). Extracto de Van den Bossche et al. (2023) traducido
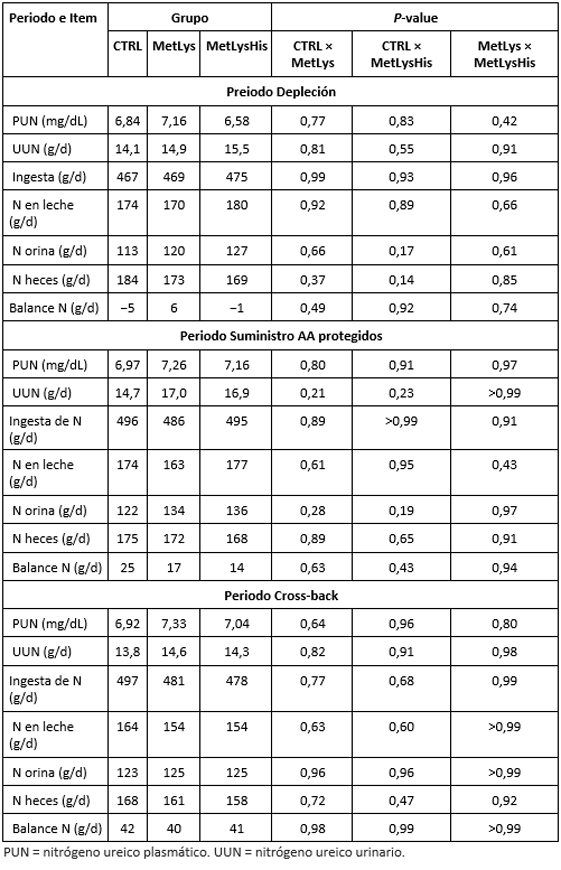
Mensajes clave:
- es importante tener en cuenta todos los EAA
- los autores observaron cómo el perfil de plasma AA indicó suministro adicional de dMet, dLys y dHis
- la suplementación de RP Met y RP-Lys o adicionalmente también RP-His a una dieta baja en proteínas no afectó el DMI, el rendimiento de la leche o MNE, lo cual, tal y como afirman los autores, se debiera a una deficiencia de Met y Lys, a pesar de la suplementación, debido a una sobreestimación del suministro de dAA en la dieta control
- este desequilibrio disminuye la eficiencia de la utilización de AA para la síntesis de proteínas de la leche, lo cual, pudo observare en una mayor concentración de MUN y numéricamente aumento de la excreción urinaria de N cuando RP-Met, RP-Lys, y RP-His se agregaron a la dieta baja en proteínas. Esto indica un aumento del catabolismo del exceso de His.
3. LA FUNCIONALIDAD DE LOS AMINOACIDOS: Lisina y su rol en inmunidad, metabolismo energético, inflamación y periodo de transición Fuente: Feeding rumen-protected lysine altered immune and metabolic biomarkers in dairy cows during the transition period. L. K. Fehlberg et al. (2023)
En línea con esta importancia de cuidar el aporte de los aminoácidos también individualmente, es necesario prestar atención a la posible funcionalidad de los mismos. En este sentido, existe cierto desconocimiento acerca del rol de la lisina en aspectos como la inmunidad y metabolismo energético.
3.1 Inmunidad
El efecto de los aminoácidos sobre la función inmunológica no debe menospreciarse, ya que se ha descrito un balance proteico negativo debido a la demanda de crecimiento del tejido mamario y fetal, lo que puede resultar en un deterioro de la función inmunológica y del tejido hepático (Connell et al., 1997; Bell et al., 2000).
En este sentido, Fehlberg et al. (2023) evaluaron el efecto de la alimentación con lisina protgidA en preparto, posparto, y ambos, en vacas Holstein multíparas, sobre el estado inmunometabólico basado en metabolitos sanguíneos, expresión de ARNm y proteínas, composición del tejido hepático y función de neutrófilos y monocitos.
- Periodo: −26 ± 4,6 días antes del parto (0,54 % RPL de ingesta de materia seca en la dieta) hasta 28 días posparto (0.39% RPL de consumo de materia seca en la dieta)
- Animales: 65 cinco vacas Holstein multíparas, por paridad, producción de leche, fecha esperada de parto y CC durante el período seco
- Grupos/tratamientos: asignadas al azar, en un diseño factorial 2 × 2.
- Preparto:
- PRE-L: preparto, TMR + lisina protegida (RPL)
- PRE-C: preparto, TMR sin RPL (PRE-C),
- Pospasrto:
- TMR + PRE-L preparto y posparto
- TMR + PRE-L preparto y PRE-C posparto
- TMR + PRE-C preparto y PRE-L posparto
- TMR + PRE-C preparto y posparto en 300 g de melaza.
- Parámetros: se tomaron muestras de sangre el día −7 ± 0,5, 0 ± 0,5, 7 ± 0,9, 14 ± 0,9 y 28 ± 0,5 en relación con el parto. Se tomaron muestras de sangre completa en −14 ± 0,5, −7 ± 0,5, 7 ± 0,9 y 14 ± 0,9 días en relación con el parto para el estallido oxidativo y la capacidad fagocítica de monocitos y neutrófilos. Las muestras de hígado se recolectaron a través de una biopsia en −12 ± 4.95 y 13 ± 2.62 d en relación con el parto y se analizaron para determinar la composición del hígado (concentraciones de triacilglicéridos y carnitina), la expresión de ARNm de genes hepáticos y la abundancia de proteínas. La abundancia de proteínas se calculó normalizando las bandas de intensidad para una proteína específica con gliceraldehído-3-fosfato deshidrogenasa.
A modo de conclusiones, el trabajo mostró que las vacas que consumieron RPL durante el período de transición tuvieron cambios a nivel de composición y funcionalidad hepáticas, función hepática mejorada (mayores concentraciones de albúmina y proteína total en plasma), y estado inmunológico mejorado (indicado por haptoglobina disminuida, actividad de glutatión peroxidasa y ARNm).
3.2 Metabolismo energético, inflamación y transición
En lo que respecta a la inflamación, según Fehlberg et al. (2023), es probable que el equilibrio adecuado de IAA durante el período de transición mejore los biomarcadores plasmáticos y la función hepática. Por ejemplo, cuando las vacas consumieron RPM durante el período de transición, hubo una disminución de las concentraciones de haptoglobina plasmática y un aumento de la fagocitosis de neutrófilos y monocitos ex vivo y de la actividad de explosión oxidativa (Zhou et al., 2016).
Figura 2. Interés de la lisina en el metabolismo energético (periodo de transición). Fuente: elaboración propia a partir de Fehlberg et al. (2023)

El estrés oxidativo, caracterizado por un aumento en la producción de especies reactivas de oxígeno, también puede contribuir sustancialmente al ambiente inflamatorio durante el período de transición (Sordillo et al., 2009). Los aminoácidos tienen la capacidad de ayudar a reducir el estrés oxidativo y ayudar con la inflamación (Li et al., 2007; Coleman et al., 2020). Por ejemplo, RPM aumentó el glutatión en el hígado (Osorio et al., 2014) y las concentraciones de albúmina plasmática mientras disminuía las concentraciones de haptoglobina plasmática durante el período de transición (Zhou et al., 2016).
A modo de conclusión de este punto 3, cabe destacar las conclusiones de Fehlberg et al. (2023), quienes determinaron, a nivel general que, las vacas que consumieron RPL probablemente pudieron controlar la activación inmunológica mientras mantenían la función hepática; y en particular, que:
- las vacas que consumieron RPL durante el período de transición, mostraron resultados mejorados en biomarcadores relacionados con la función hepática y un estado inflamatorio disminuido;
- su tendencia a aumentar las concentraciones de proteínas de fase aguda negativa probablemente indica una función hepática mejorada, lo que,
- junto con la disminución de las concentraciones de haptoglobina en la sangre y la disminución de la expresión del ARNm hepático de SAA3 posparto, cuando se consumió RPL antes del parto en comparación con cuando no se consumió RPL, puede indicar una disminución de la inflamación. En conclusión,.
Publicado originalmente en Entorno Ganadero n120
Temas relacionados:
Preguntas Relacionadas
¿Qué efecto tienen los aminoácidos sobre la función inmunológica durante el período de transición?
El efecto de los aminoácidos sobre la función inmunológica no debe menospreciarse, ya que se ha descrito un balance proteico negativo debido a la demanda de crecimiento del tejido mamario y fetal, lo que puede resultar en un deterioro de la función inmunológica y del tejido hepático.
Autores:

Orffa Excentials
Referentes que Recomendaron :
Umberto Francesa, Alfredo J. EscribanoRecomendar
Comentar
Compartir

28 de julio de 2023
En la practica, uno de los problemas que vacas en alta producción manifiestan con dietas bajas en proteínas, es consumo, el cual se reduce lo suficiente para causar problemas en producción de leche.
No cabe duda que el control en la elimination de nutrientes y gases invernadero al medio ambiente, va a permanecer el tema de discusión durante muchos anos, y francamente una amenaza legal para la lechería comercial en el futuro. En los EEUU, por ejemplo, algunas municipalidades se han adelantado al gobierno federal en crear leyes locales que prohíben cierto tipo de industrias dentro de esas municipalidades, incluyendo ciertos tipos de ganaderías.
Interesante la participación de Lisina en la respuesta inmunitaria de tipo celular.
Recomendar
Responder
28 de julio de 2023
Gracias Alfredo por tomarse el tiempo en la traducción de este documento.
Recomendar
Responder

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar

Eurotec Nutrition Argentina S.R.L.