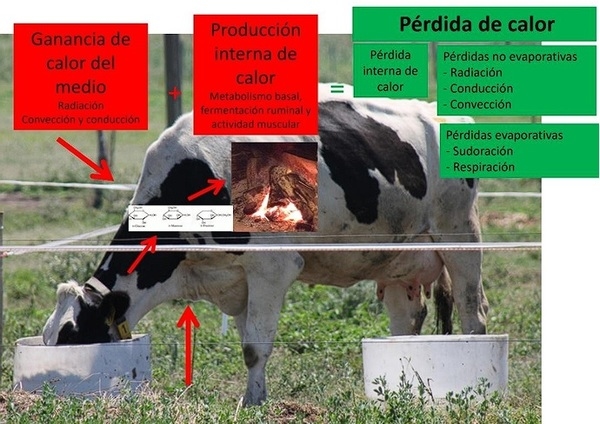
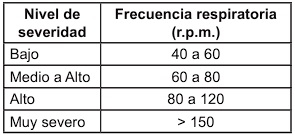
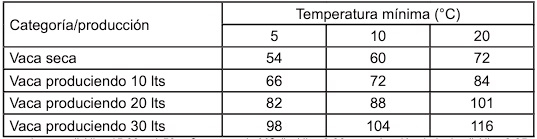
Efecto del estrés calórico en el animal
Adin, G.; Gelman, A.; Solomon, R.; Fla menbaum, I.; Nikbachat, M.; Yosef, E.; Zenou, A.; Shamay, A.; Feuermann, Y.; Mabjeesh, S.J.; Miron, J. 2009. Effects of cooling dry cows under heat load conditions on mammary gland enzymatic activity, intake of food and water, and performance during the dry period and after parturition. Livestock Science 124 189–195
Avendano-Reyes, L., F. Alvarez-Valen zuela, D.; Correa-Calderon, A.; Saucedo Quintero, J. S.; Robinson, P. H.; Fadel, J. G. 2006. Effect of cooling Holstein cows du ring the dry period on postpartum performan ce under heat stress conditions. Livest. Sci. 281:2535–2547.
Baker CC, Coppock CE, Lanham JK, Nave DH, Labore JM, Brasington CF, Stermer RA. 1988. Chilled drinking water effects on lactating Holstein cows in summer. Journal of Dairy Science. 71 (10): 2699-2708.
Baumgard L, Rhoads RP. 2007. The Effects of Heat Stress on Nutritional and Manage ment Decisions. [en línea]. 15 agosto 2012. http://www.wdmc.org/2009/The%20Effects%20 of%20Het%20Stress%20on%20Nutritional%20 &%20Management%20Decisions.pdf
Baumgard L, Rhoads RP. 2012. Effects of Environment on metabolism. En: Collier J, Collier JL. (Eds.). Environment physiology of Livestock. Iowa: John Wiley and Sons, Inc. 81-100.
Beede DK, Collier RJ. 1986. Potential nu tritional strategies for intensively managed cattle during thermal stress. Journal of Ani mal Science. 62: 543-554.
Berman A, Folman Y, Kaim M, Mamen M, Herz Z, Wolfenson D, Arieli A, Graber Y. 1985. Upper critical temperatures and for ced ventilation effects for high-yielding dairy cows in a subtropical climate. Journal of Dairy Science. 68: 1488-1495.
Bernabucci U, Bani P, Ronchi B, Lacetera N, Nardone A. 1999. Influence of short and long-term exposure to a hot environmental rumen passage rate and diet digestibility by Friesian heifers. Journal of Dairy Science. 82: 967–973.
Bernabucci U. 2012. Impact of the hot envi roment on nutrient requeriments. En: Collier J, Collier JL. (Eds.). Enviroment Phisiology of Livestock. Iowa: Jhon Wiley and Sons, Inc. 101-128 p
Bouraqui R, Lahmar M, Majdoub A, Dje mali M, Belyea R. 2002. The relationship of temperature-humedity index with production of dairy cows in a Mediterraneum climate. Animal Research. 51: 479-491.
Breinholt KA, Gowen FA, Nwosu CC. 1981. Influence of environment and animal factor son day and night grazing activity of imported Holstein-Friesian cows in the humid lowland tropics of Nigeria. Tropical Animal Produc tion. 6 (4): 300-307
Brown-Brandl TM, Eigenberg RA, Niena ber JA, Hahn GL. 2005a. Dynamic response indicators of heat stress in shaded and non shaded feedlot cattle, Part 1: Analyses of indicators. Biosystems Engineering. 90 (4): 451–462.
Collier RJ, Beede DK, Thatcher WW, Is rael LA, Wilcox CJ. 1982. Influences of en vironment and its modification on dairy ani mal health and production. Journal of Dairy Science. 65 (11): 2213-2227.
Collier RJ, Eley RM, Sharma AK, Pereira RM, Buffington DE. 1981. Shade manage ment in subtropical environment for milk yield and composition in Holstein and Jersey cows. Journal of Dairy Science. 64: 844-849.
Collier, R. J.; Doelger, S. G.; Head, H. H.; Thatcher, W. W.; Wilcox, C. J. 1982 b. Effects of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and postpartum milk yield of Holstein cows. J. Anim. Sci. 54:309–319.
Do Amaral, B. C.; Connor, E. E.; Tao, S.; Hayen, J.; Bubolz, J.; Dahl, G. E. 2009. Heat-stress abatement during the dry period: Does cooling improve transition into lacta tion? J. Dairy Sci. 92 :5988–5999
Dowling DF. 1955. The hair follicle and apo crine gland populations of Zebu (Bos indicus L.) and Shorthorn (B. taurus L.) cattle skin. Australian Journal of Agricultural Research. 6 (4): 645–654.
Ferreira F, Campos WE, Carvalho AU, Pires MF, Martinez ML, Silva MV, Verneque RS, Silva PF. 2009. Taxa de sudação e parâme tros histológicos de bovinos submetidos ao estresse calórico. Arquivo Brasileiro de Medi cina Veterinária e Zootecnia. 61 (4): 763-768.
Findlay JD, Yang SH. 1950. The sweat glands of Ayrshire cattle. Journal of Agricul tural Science. 40: 126-133.
Frazzi E, Calamari L, Calegari F, Stefanini L. 2000. Behavior of dairy cows in response to different barn cooling systems. Transac tion of the American Society of Agricultural Engineers. 43 (2): 387-394.
Gallardo M, Valtorta S. 2011. Producción y bienestar animal. Estrés por calor en ganado lechero: impactos y mitigación. Editorial He misferio Sur S.A. Argentina. 128 pp.
Gebremedhin KG. 2012. Impact of the hot enviroment on nutrient requeriments. En: Collier J, Collier JL. (Eds.). Enviroment Phi siology of Livestock. Iowa: Jhon Wiley and Sons, Inc. 35-48.
Hansen PJ. 2004. Physiological and cellular adaptations of zebu cattle to thermal stress. Animal Reproduction Science. 82 (83): 349 360.
Johnson HD. 1987. Bioclimatology and the Adaptation of Livestock. En: Johnson HD (Eds.). Bioclimatology and adaptation of Li vestock. Amsterdam. Elsevier Netherlands. 279.
Kadzere CT, Murphy MR, Silanikove N, Maltz E. 2002. Heat stress in lactating dairy cows: A review. Livestock Production Scien ce. 77: 59-91.
Lanham JK, Coppock CE, Milam KZ, Labo re JM, Nave DH, Stermer RA, Brasington CF. 1986. Effects of drinking water tempera ture on physiological responses of lactating Holstein cows in summer. Journal of Dairy Science. 69: 1004–1012.
McDowell RE, Hooven NW, Camoens JK. 1976. Effect of climate on performance of Holsteins in first lactation. Journal of Dairy Science. 59 (5): 965–973.
Milam KZ, Coppock CE, West JW, Lanham JK, Nave DH, Labore JM, Stermer RA, Bra sington CF. 1986. Effects of drinking water temperature on production responses in lac tating Holstein cows in summer. Journal of Dairy Science. 96: 1013-1019.
Mishra M, Martz FA, Sranleu RW, Johnson HD, Campbell JR, Hilderbrand E. 1970. Effect of diet and ambient temperature–hu midity on ruminal pH, oxidation reduction po tential, ammonia and lactic acid in lactating cows. Journal of Animal Science. 30: 1023 1028.
Moody EG, Van-Soest PJ, McDowell RE, Ford GL. 1967. Effect of high temperature and dietary fat on performance of lactating cows. Journal of Dairy Science. 50 (12): 1909-1916.
Muller CJ, Botha JA. 1994. Effect of shade on various parameters of Friesian cows in a Mediterranean climate in South Africa. 1. Feed and water intake, milk production and milk composition South African Journal of Animal Science. 24 (2): 47-55.
Murphy, M. R., C. L. Davis, and G. C. Mc Coy. 1983. Factors affecting water consump tion by Holstein cows in early lactation. J. Dairy Sci. 66:35–38.
Nardone, A.; Lacetera, N.; Bernabucci, U.; Ronchi, B. 1997. Composition of Co lostrum from Dairy Heifers Exposed to High Air Temperatures During Late Pregnancy and the Early Postpartum Period. J Dairy Sci 80:838–844.
NRC (National Research Council). 2001. Nutrient requirements of dairy cattle. Seven th revised edition. National Academic Press, Washington DC.
O’Brien MD, Rhoads RP, Sanders SR, Duff GC, Baumgard LH. 2010. Metabolic adapta tions to heat stress in growing cattle. Domes tic Animal Endocrinology. 38: 86–94.
O’Brien MD, Cole LC, Wheelock JB, San ders SR, Duff GC, Baumgard LH, Rhoads RP. 2008. Thermal and nutritional regulation of hepatic gluconeogenic genes in growing beef cattle. Journal of Animal Science 86. (2): 455-467.
Pocay PL, Pocay VG, Starling JM, Silva RG. 2001. Respostas fisiológicas de vacas Holandesas predominantemente negras sob radiaçao solar directa. Ars Veterinaria. 17(2): 155-161.
Purwanto BP, Abo Y, Sakamoto R, Furu moto F, Yamamoto S. 1990. Diurnal pat terns of heat production and heart rate under thermoneutral conditions in Holstein Friesian cows differing in milk production. The Journal of Agricultural Science. 114: 139-142.
Ravagnolo O, Misztal I, Hoogenboom G. 2000. Genetic component of heat stress in dairy cattle, development of heat index function. Journal of Dairy Science. 83: 2120 2125
Regan WM, Richardson GA. 1938. Reac tions of the dairy cows changes in environ mental temperature. Journal of Dairy Scien ce. 21: 73-79.
Rhoads ML, Rhoads RP, Vanbaale MJ, Collier RJ, Sanders SR, Weber WJ, Cro oker BA, Baumgard LH. 2009. Effects of heat stress and plane of nutrition on lactating Holstein cows: I Production, metabolism, and aspects of circulating somatotropin. Journal of Dairy Science. 92: 1986–1997.
Saravia C. 2009. Efecto del estrés calórico sobre las respuestas fisiológicas y produc tivas de vacas Holando y Jersey. Tesis de Maestría en Ciencias Agrarias. Montevideo, Uruguay. Facultad de Agronomía. 140 p.
Seath DM, Miller GD. 1946. Effect of warm weather on grazing performance of milking cows. Journal of Dairy Science. 29: 199 206.
Shwartz G, Rhoads ML, Vanbaale MJ, Rhoads RP, Baumgard LH. 2009. Effects of a supplemental yeast culture on heat-stres sed lactating Holstein cows. Journal of Dairy Science. 92: 935–942.
Spiers D.E. 2012. Physiological basics of temperatura regulation in domestic animals. En: Collier J, Collier JL. (Eds.). Environment Physiology of Livestock. Iowa: John Wiley and Sons, Inc. 17-34.
Spiers DE, Spain JN, Sampson JD, Rhoads RP. 2004. Use of physiological parameters to predict milk yield and feed intake in heat stressedd airy cows. Journal of Thermal Bio logy. 29: 759–764.
Tapki I, Sahin A. 2006. Comparison of the thermoregulatory behaviours of low and high producing dairy cows in a hot environment. Animal Behaviour Science. 99: 1–11.
Thomas CK, Pearson RA. 1986. Effects of ambient temperature and head cooling on energy expenditure, food intake and heat to lerance of Brahman and Brahman x Friesian cattle working on treadmills. Animal Produc tion. 43: 83-90.
Urdaz, J. H.; Overton, M. W.; Moore, D. A.; Santos, J. E. P. 2006. Technical Note: Effects of Adding Shade and Fans to a Fee dbunk Sprinkler System for Preparturient Cows on Health and Performance. J. Dairy Sci. 89:2000–2006.
Valtorta SE, Leva PE, Gallardo MR, For nasero LV, Veles MA, Garcia MS. 1997. Producción de leche: respuestas a la alta temperatura. Archivo Latinoamericano de Producción Animal. 5 (1): 399-401.
West JW. 2003. Effects of heat-stress on production in dairy cattle. Journal of Dairy Science. 86: 2131–2144.
West JW. 2003. Effects of heat-stress on production in dairy cattle. Journal of Dairy Science. 86: 2131–2144.
Widowski TM. 2001. Shade-seeking beha vior of rotationally-grazed cows and calves in a moderate climate. En: International Sym posium of Livestock Environment (6º, 2001, Louisville, Kentucky). International Sympo sium of Livestock Environment. Ray Bucklin and Robert W. Bottcher. pp 632-639.
Wolfenson, D., Flamembaum, I. Berman, A. 1988. Dry period heat stress relief effects on prepartum progesterone, calf birth weight, and milk production. J. Dairy Sci. 71:809 818.
Yousef MK. 1985. Stress physiology: defini tion and terminology. En: Yousef MK. (Eds.). Stress physiology in livestock. Florida: Boca Raton. 3-7.







")



