Historia.
El rumen de los rumiantes es una maravilla evolucionaria que les ha permitido sobrevivir y multiplicarse durante miles de años. Es el perfecto ejemplo de un ecosistema complejo entre un organismo superior en simbiosis con otros organismos inferiores beneficiándose mutuamente.
La mayoría de los especies incluyendo el hombre, poseen sistemas simbióticos en una forma u otra; sin embargo, son los rumiantes los que lo han perfeccionado al máximo de eficiencia esa relación. Los rumiantes poseen la capacidad de lograr sobrevivir bajo tremendas condiciones de escasez y hasta llegar a reproducirse bajo esas condiciones.
Recordemos que nuestros rumiantes domésticos, aunque con algunas variantes de manejo, pueden perfectamente vivir exitosamente en condiciones naturales semejantes a aquellos rumiantes salvajes que hoy pastorean sobre praderas naturales.
El estudio de este sistema ecológico es incluido en el currículo de estudiantes iniciándose en cualquier carrera que incluye las ciencias biológicas (Hungate, 1966).
Aristóteles describió la relación entre los molares del bovino, la dieta fibrosa y la complejidad de los estómagos del rumiante en la manera siguiente:
“cuando un animal es de gran tamaño y se alimenta de sustancias duras y lignosas en carácter…podría como consecuencia tener varios estómagos” (Lennox, 2001).
Introducción.
Los rumiantes han desarrollado un diverso y sofisticado sistema ecológico para la digestión de forrajes fibrosos. Las paredes celulares de las plantas consisten de una estructura química compleja y no completamente dilucidada.
La digestibilidad de la fibra en forrajes de clima tropical (C4) difiere de aquellos forrajes en climas templados (C3). En áreas más calientes, forrajes C3 no prosperan durante el verano, siendo esta una particularidad de especies; pastos C3 crecen mejor durante la primavera y el otoño, permaneciendo semidormidos durante los meses calientes del verano.
Los pastos tropicales C4 crecen durante todo el año y contienen un gran número de especies que difieren en su adaptabilidad, reproducción y valor nutritivo. Pastos de clima templado C3, pueden perfectamente adaptarse a las montañas altas tropicales arriba de los 1600 msnm, lo que ofrece una ventaja económica que discutiremos más adelante.
Como veremos más adelante, una de las diferencias entre forrajes C3 y forrajes C4, es la digestibilidad de los componentes en sus paredes celulares lo que determina entre estos dos grupos tremendas diferencias en su valor nutritivo.
Diferencias entre los Pastos Tropicales (C4) y Meridionales (C3).
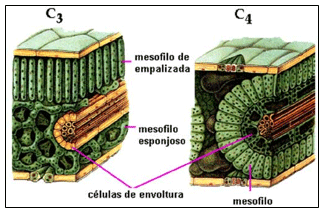
Las diferencias anatómicas entre pastos del tipo Panicoide (tropical) y tipo Festucoide (meridional), están asociadas al proceso de fotosíntesis que estos llevan a cabo (Wilson and Hacke, 1987). Ellos observaron que las hojas de pastos tropicales C4, contenían en su estructura un número menor de células “mesófilas” y un mayor número de células epidérmicas, tejido vascular y esclerénquima; en los cuales, el polisacárido celulosa tiende a acumularse conforme la planta avanza en edad. Esta situación fisiológica confiere a los pastos tropicales una menor digestibilidad cuando se comparan con aquellos pastos de clima templado.
Comparaciones de pastos del tipo panicoide (C4) y festucoide (C3), bajo similares condiciones de temperatura y luz, demuestra que son las diferencias morfológicas en sus hojas las que claramente contribuyen a la baja digestibilidad de la materia seca de los pastos tropicales (Akin et al., 1983).
La figura 1 muestra un arreglo de los diferentes tejidos y composición celular en las hojas de pastos C3 y C4.
Análisis cuantitativo de la anatomía de las hojas en diferentes pastos de tipo tropical, demostró un 25% menos digestibilidad en sus tejidos que aquellos analizados para pastos de climas templados (Akin and Burdick, 1975).
Claramente, la energía contenida en los forrajes C4 es limitada por la barrera física de fibra en la pared celular.
La proporción de fibra y carbohidratos solubles es un determinante de importancia en la energía neta disponible en un forraje. Rumiantes, solamente digieren entre un 40 a un 70% de la fibra diaria ingerida.
Lignina interfiere con la degradación de los polisacáridos en la fibra, actuando como una barrera física y creando puentes ferulados entre los polisacáridos celulosa y hemicelulosa (Moore and Jung, 2001). Lignina es más concentrada en pastos tropicales (C4) que en pastos C3 y es la base química para la baja digestibilidad de los pastos tropicales (Ramalho, 1991).
Lignina y la Digestión de la Fibra.
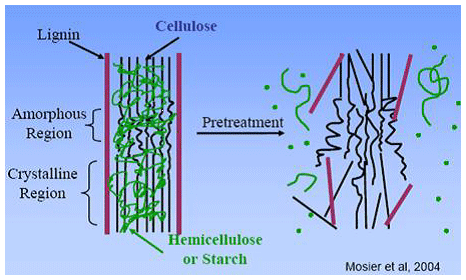
La lignina es un polímero formado de monolignoles derivados de la vía fenilpropanoide de las plantas vasculares. Se deposita en las paredes celulares y se incrementa durante el proceso de maduración de la planta. En los forrajes, la lignina se considera como un componente anti-cualitativo por su impacto negativo en la disponibilidad nutricional de la Fibra detergente Neutra (FDN); así como, de aquellos nutrientes orgánicos el contenido interior de las células. La lignina interfiere con la digestión de los polisacáridos de la pared celular al actuar como barrera física para las enzimas microbianas.
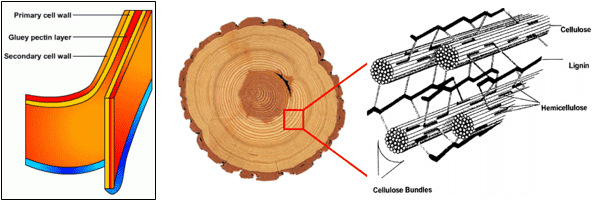
Figura 2.- Esta figura muestra un ejemplo de un tratamiento físico-químico industrial para romper la lignina. El rumen de los bovinos, no posee la capacidad de romper las paredes celulares en semejante manera.
La lignificación tiende a ser más intensa en tejidos estructurales como el xilema y esclerénquima. Los órganos de la planta que contienen altas proporciones de estos tejidos, tales como los tallos, son menos digestibles que aquellos que contienen bajas concentraciones. La proporción de tejidos y órganos lignificados típicamente aumenta conforme la planta madura, por lo que a menudo hay una relación negativa entre la digestibilidad y madurez.
La temperatura, humedad del suelo, luz y fertilidad del suelo pueden tener también efectos directos o indirectos en la lignificación. Las prácticas de manejo más útiles para minimizar los efectos negativos de la lignificación son la manipulación de las comunidades vegetales para que contengan más especies deseables, y el manejo de forrajes al estado vegetativo ideal.
La lignificación controla la cantidad de fibra que puede ser digerida y por lo tanto, tiene un impacto directo sobre la “energía digestible (ED)” de un forraje (Jung and Allen 1995).
La lignificación también influencia la cantidad de materia seca (MS) que el animal puede consumir (Mertens, 1994). La porción no digerida del forraje pasa lentamente a través del retículo-rumen y contribuye al llenado del rumen (ruminal fill), efecto que reduce el consumo de materia seca (sensación de llenado).
Entre mayor es la concentración de fibra no degradable en la dieta, el consumo de la misma es reducido; por lo tanto, la lignificación impacta el valor nutritivo del forraje de dos maneras:
a) reduciendo la concentración de energía digestible (ED) y
b) limitando el consumo de materia seca (Moore et al. 1993).
Lignina y calidad del forraje.-
Está bien establecido como un hecho que la concentración de lignina en los forrajes es negativamente correlacionada con su digestibilidad (Jung and Deetz, 1993).
La razón para esta correlación es porque la concentración de lignina siempre incrementa conforme la concentración de la pared celular aumenta, y la pared celular de los forrajes son siempre menos digestibles que los componentes celulares solubles en el interior de la célula.
En general, la concentración de lignina en leguminosas es menor que en pastos, sugiriendo que la lignina en las paredes celulares de los pastos ofrece una pared inhibitoria más poderosa a la acción de las enzimas contenidas en la microflora ruminal (Van Soest 1964, Buxton and Russell 1988). Esta conclusión hay que tomarla con cuidado, ya que se ha tomado de investigaciones hechas utilizando métodos de análisis en donde la lignina es expuesta a ácidos detergentes (ADL). Este método ha probado no ser confiable en sus estimaciones con respecto a las concentraciones de lignina en pastos. Estudios microscópicos han demostrado que lo opuesto es más probable y que la lignina es más inhibitoria en leguminosas que en pastos, ya que la lignina en legumbres es completamente indigestible, mientras que las paredes celulares de los pastos son digeridas en cierto grado (Engels 1989, Engels and Jung 1998).
Varios mecanismos han sido sugeridos para explicar cómo la lignina inhibe la digestión de la pared celular; sin embargo, se ha llegado a la conclusión general que la lignina simplemente actúa como una barrera física a las enzimas de los microbios del rumen, previniendo que alcancen los polisacáridos contenidos dentro de las paredes celulares (Chesson 1993, Jung and Deetz 1993).
Lignina en mutantes bm.-
Fue propuesta la noción que la composición de la lignina, medida como el radio syringyl a guaiacyl, alteraría el impacto inhibitorio sobre la digestión de la pared celular de bm mutantes porque estos tienen una mejor digestibilidad de las paredes celulares y por lo tanto, una menor cantidad de lignina del tipo syringyl. Sin embargo, los bm mutantes también tienen una menor cantidad de lignina total y una menor concentración de lignina en las paredes celulares. Los mutantes bm no están dotados para probar la validez de la hipótesis de que es la composición química de la lignina la que afecta la digestibilidad de la misma (Jung and Deetz 1993).
Puentes Ferulados de lignina en las paredes celulares de pastos.-
Jung and Deetz (1993), también propusieron que son los puentes ferulados de lignina en los polisacáridos de la pared celular en los pastos, los que impiden la digestión celular. Intentos en demostrar esta teoría han sido inconclusos debido a que los puentes ferulados comienzan a formarse muy temprano durante el crecimiento de las plantas, y subsecuente deposición de lignina sobre los polisacáridos de la pared celular, irían a diluir su concentración (Morrison et al. 1998). Sin embargo, utilizando un modelo con cultivos de maíz, Grabber et al. (1998), demostraron que reduciendo los puentes ferulados se incrementaba la digestibilidad de la pared celular.
Casler y Jung (1999) demostraron que algunos genotipos del pasto Bromus innermis Leyss, seleccionados por tener menores concentraciones de puentes ferulados, exhibieron mejores digestibilidades de las paredes celulares. Es muy probable entonces, que son los puentes ferulados de lignina embebidos en los polisacáridos de la pared celular los que le confieren su digestibilidad.
Factores afectando la lignificación.
Existen un gran número de factores que afectan la calidad de los forrajes y que pueden influenciar el valor nutritivo de los mismos.
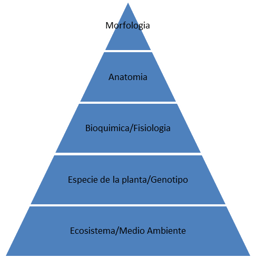
Estos factores pueden ser arreglados en un orden jerárquico que indican la relativa dependencia entre los mismos
Figura 3.
Todos estos procesos son dinámicos y tienen mecanismos de retroalimentación continua entre los diferentes niveles. Consecuentemente, existen un gran número de interacciones entre ellos que pueden ocurrir, las cuales hacen más difícil el dilucidar sus efectos en el valor nutricional de los forrajes.
Los efectos de la lignina en la calidad del forraje pueden ser alterados y por tanto manipulados en cualquier nivel de la jerarquía de los factores afectando el valor nutritivo de un cultivo de forrajes de una especie determinada.
Ecosistema/Medio Ambiente.-
Este es el nivel que más en cuenta habría que considerar al decidir cuál o cuáles serían los forrajes más adaptables a un área geográfica específica. Factores edafológicos del suelo, geográficos, topografía del suelo, climáticos, y bióticos (pestes y enfermedades), asociados con ese medio ambiente determinarían el potencial con que ciertas especies de forrajes podrían ser cultivadas ahí. La variabilidad y espectro entre estos ecosistemas es amplio. Algunos de ellos reúnen las condiciones ambientales y geográficas en donde se podrían cultivar un gran número de diferentes cultivos de alta productividad; mientras que otros, son más exigentes y demandan mayor riesgo.
Manipulación es posible, mediante la utilización de tecnología agraria; tales como: fertilidad del suelo, riego artificial, control de pestes, cultivo en bandas, etc.
En contraste, hay diversos y extremos sistemas ecológicos que son menos productivos. Estos ecosistemas no son fácilmente manipulados y consecuentemente tienen un potencial limitado por las condiciones existentes de la zona. Las modificaciones que el manejo podría provocar son limitadas y económicamente costosas. La producción de forraje en estas áreas generalmente involucra el manejo de plantas nativas a través de pastoreo estacional en donde los rebaños son movidos continuamente de un lugar a otro siguiendo el ciclo vegetativo de las plantas. El mejor ejemplo seria el tipo de ganadería tradicional todavía utilizado en Sub-Sahara África por tribus de ganaderos nómadas.
Especies de Forrajes/Genotipo.-
Como mencionábamos antes, comunidades de forrajes pueden ser modernas o nativas y su establecimiento depende de los factores del medio ambiente descritos anteriormente.
La concentración y composición de la lignina es muy variable entre las diferentes especies y géneros. La mayoría de los forrajes modernos cultivados hoy en día pertenecen; ya sea, a la familia de los pastos o Poaceae (monocotiledóneas) o a la familia de las leguminosas o Fabáceas (dicotiledóneas). Existen grandes diferencias en lignificación entre los pastos y las leguminosas y su impacto en el valor nutritivo del forraje.
La concentración de lignina en pastos y leguminosas es muy similar cuando expresada en términos de su contenido en materia seca (MS). Sin embargo, cuando es expresada en la proporción incluida en la “fibra detergente neutra” (FDN), las leguminosas han demostrado una cantidad más amplia en la concentración de lignina y es generalmente más alta que la de los pastos (Buxton and Brasche 1991, Buxton and Russell 1988).
La amplitud en la digestibilidad de la fibra es similar entre pastos y leguminosas, indicando que hay diferencias entre las dos familias en la manera que la lignina interacciona con los otros componentes de la fibra (celulosa y hemicelulosa) (Buxton et al. 1987, Buxton 1989).
La principal diferencia entre pastos y leguminosas radica en la concentración de fibra. Los pastos tienen una mayor concentración de fibra que las leguminosas, lo que los hace tener una menor cantidad de contenidos nutricionales intracelulares (Buxton and Russell 1988). De tal manera, aunque la lignina impacta negativamente la digestibilidad de las leguminosas, las leguminosas tienen una menor cantidad de fibra Cruda (FDN) que los pastos, lo que afecta directamente el valor energético entre ambos. Por esta razón, la concentración de lignina no es un buen indicador de energía digestible (ED), al hacer comparaciones entre pastos y leguminosas. En las leguminosas las hojas son altamente digestibles y serían los tallos los que contienen paredes altamente lignificadas.
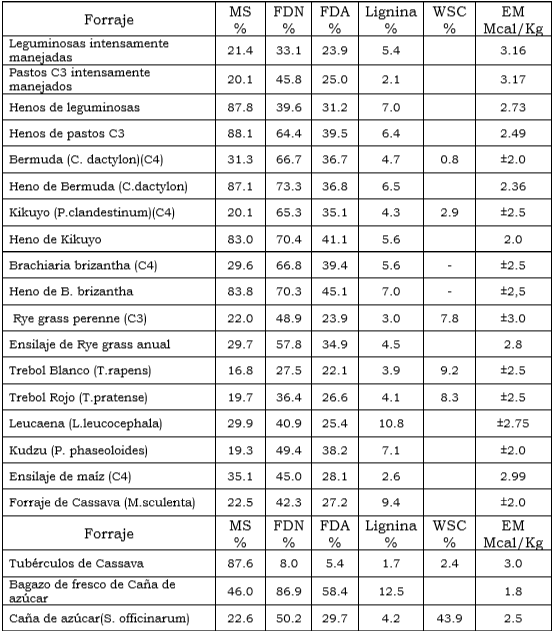
Tabla 1. Contenidos en Fibra de algunas Leguminosas y Pastos de altura.
Pastos de clima templado y pastos tropicales poseen una composición similar en fibra y lignina (Jung and Vogel 1986, 1992). La gran diferencia es establecida por la mayor acumulación de fibra en los pastos tropicales, lo que los hace tener una concentración mayor de lignina en estados de crecimiento vegetativo similares.
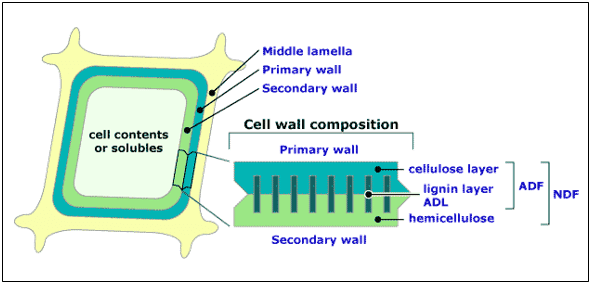
Figura 2.- La lignificación ocurre principalmente en la pared celular primaria o externa limitando con la lamella intercelular.
Aun dentro de especies; individuos o cultivars, presentan significantes diferencias en lignificación como resultado de manipulación genética. Estas diferencias proveen la variabilidad genética necesaria para hacer mejoramientos en la calidad nutritiva de un forraje determinado a través de métodos tradicionales de cruzamiento. Selección con esos métodos en alfalfa (Medicago sativa L), han creado variedades con cantidades menores de lignina; debido a plantas con mayor cantidad de hojas y cortos tallos, con lo que se reduce el porcentaje de lignina en la FDN. Con esta manipulación genética en la morfología, la fibra fue drásticamente disminuida.
Seleccionando por digestibilidad en pastos, ha sido relacionada en algunos casos a cambios en lignificación, un ejemplo es en el pasto “smoothbrome grass”, en donde lignificación ha sido reducida a través de selección de mejores cultivars. En otros casos, se ha mejorado la digestibilidad de un pasto sin haber cambiado la concentración de la lignina, sino la manera en que la lignina interacciona con los otros carbohidratos de la pared celular, como en el caso de “switch grass”.
Bioquímica / Fisiología.-
Diferencias genotípicas entre los forrajes son expresadas a un nivel bioquímico,
como respuesta al estímulo del medio ambiente. La síntesis de lignina es un proceso complejo que involucra un número de reacciones bioquímicas mediadas por enzimas (Boudet 1998). Alterando estas enzimas, iría a cambiar la concentración de lignina en los forrajes. Un buen ejemplo es con bm mutantes. Esta mutación es caracterizada por una coloración oscura de la vena media de la hoja, y es usualmente relacionada a una más baja concentración de lignina en la FDN, la cual es también alterada en su composición (Cherney et al 1991). Forrajes de este mutante (maíces BMR), are potencialmente más digestibles que sus contrapartes debido a que muestran los atributos antes mencionados. Esta característica puede ser inducida por muta génesis y es de esa manera que se han obtenido sorgos bm (Sorghum bicolor).
Los efectos fenotípicos de mutaciones bm han sido bien estudiados y hoy sabemos con certidumbre que su establecimiento se debe a alteraciones en la actividad enzimática en la composición de la lignina. El rasgo bm es de particular interés en forrajes debido a su asociación en la digestibilidad de la FDN; sin embargo, debido a diferentes factores que incluyen la opinión pública, forrajes obtenidos por medios transgénicos no ha permitido su desarrollo comercial y es muy probable que no lo vaya a tener durante un futuro cercano.
Mencionamos al principio la interacción que existe entre los diferentes niveles de factores que afectan lignificación en los forrajes (figura 2) y respuestas fisiológicas a varios factores ambientales que pueden afectar el grado de lignificación en un forraje determinado.
Temperatura, humedad del suelo, luz y fertilidad del suelo pueden tener un efecto directo o indirecto sobre la lignificación (Buxton and Casler 1993, Nelson and Moser 1994).
a) Temperatura.-
Lignificación tiende a incrementar en plantas que crecen bajos temperaturas cálidas. Esto parece ser relacionado con una mayor actividad enzimática en la síntesis de lignina o a un mayor reparto de materia seca dirigida a la formación de lignina en los tejidos. Wilson et al. (1994) descubrió que sometiendo pastos tropicales (C4) y de clima templado (C3) bajo un regimiento de alta temperatura no alteraba la proporción de sus tejidos, pero si intensificaba en ambos la lignificación y bajaba la digestibilidad de la FDN de los forrajes.
Los pastos C4 tropicales difieren en su adaptación de acuerdo a las diferentes alturas geográficas de la América Tropical y los que crecen muy bien en las sabanas calientes a nivel del mar no prosperan bien en las montañas y viceversa. Los pastos de las montañas son más reducidos en el número de especies disponibles que aquellos existentes en los climas cálidos de las sabanas, los cuales son abundantes en el número de especies disponibles. Los pastos tropicales de las montañas altas de la América Tropical, en general contienen un valor nutritivo más alto que sus contrapartes en áreas más bajas, demostrando que las altas temperaturas y alta luminosidad, influyen en la lignificación de las paredes celulares y su concentración en la FDN.
b) Humedad del suelo.-
Deficiencias de humedad en el suelo afecta la lignificación de los forrajes, aunque este efecto ocurre mayormente como función de cambios en el desarrollo morfológico de la planta (Buxton y Casler, 1993). En la alfalfa, la concentración de los componentes de la pared celular aumenta bajo irrigación artificial, pero sin afectar la composición de los mismos (Hamlin et al.1989).
Cuando agua es no es disponible en las cantidades que una especie de forraje requiere durante las diferentes etapas de su crecimiento, la digestibilidad es afectada debido a un estrés por falta de humedad. Un exceso de humedad, también afecta la deposición de lignina en los tejidos, en este caso disminuyendo la concentración de lignina y FDN en las paredes celulares (Buscaglia et al. 1994).
c) Luz.-
Intensidad y duración diurna también afectan la lignificación de los forrajes. Durante la primavera los pastos C3 de climas templados crecen vigorosamente debido a que la intensidad de la luz y la temperatura ambiental son ideales para su crecimiento. Al llegar el verano, los días son más largos y más calurosos, lo que provoca en estos pastos un cese en su crecimiento que se renueva en los meses del otoño.
Una baja intensidad en luz disminuye la lignificación pero también reduce el crecimiento de las plantas al disminuir la fotosíntesis; sin embargo, este no es el caso de los forrajes de clima templado C3, los cuales están perfectamente adaptados a condiciones de menor intensidad luminosa y la fotosíntesis se lleva a cabo a través de un método muy diferente a la de los forrajes tropicales C4, debido al mayor número de células mesófilas en sus tejidos.
El sombreado de forrajes reduce la lignificación; sin embargo, también reduce el crecimiento de los mismos. En términos generales se ha demostrado que el sombreado tiende a reducir la lignificación y consecuentemente mejora la digestibilidad de la materia seca, en forrajes tropicales C4.
La intensidad luminosa durante la época lluviosa en las montañas de la América Tropical, disminuye debido al mayor número de días nublados. Esta situación hace que forrajes de clima templado C3 se puedan adaptar perfectamente bajo estas condiciones climáticas. El pasto perenne Rye Grass Italiano es un buen ejemplo de esta adaptación. Es un pasto con una cantidad baja en lignina y con una concentración de azucares intracelulares muy aceptable en una fermentación anaeróbica, si se desea ser utilizado como pasto para ensilaje.
d) Fertilidad del suelo.-
Los nutrientes del suelo pueden tener un efecto dramático en lignificación. Deficiencias retrasan el crecimiento normal en una manera similar a la de otros factores estresantes ya revisados (Miller y Reetz, 1995). Por el contrario, existe poca evidencia que sugiera que fertilización de forrajes en suelos que ya contienen los nutrientes necesarios para el desarrollo normal de un forraje, iría a disminuir la lignificación en ese forraje o la digestibilidad de la FDN.
La respuesta en lignificación y en general la digestibilidad de la fibra bruta a la fertilización es variable entre diferentes forrajes. La fertilización con azufre ha reportado que reduce la lignificación y mejora la digestibilidad de la fibra en sorgo (Ahmad et al. 1995); sin embargo, este efecto no fue demostrado en otras especies de forrajes.
Anatomía de las Plantas.-
La lignificación ocurre durante el proceso de diferenciación celular y en general coincide con la formación de la pared celular secundaria (Iiyama et al. 1993).
La deposición de lignina es iniciada en las esquinas de la célula adyacente a la lamella y procede internamente dentro de la pared primaria y secundaria. Lignificación siempre precede a la deposición de carbohidratos estructurales en formación y detrás de la pared celular que se está en ese momento formando. Debido a esto, la concentración de lignina es generalmente mayor en la primera (exterior) pared celular; sin embargo, esto no significa que lignificación de la pared secundaria pudiese ocurrir.
Figura 3.
La concentración de lignina varía ampliamente entre células de diferentes tipos, resultando que los tejidos de la planta tienen diferentes degrabilidades en respuesta a las diferentes concentraciones en lignina. Los tejidos con altas concentraciones de lignina incluyen: la epidermis en los pastos, tejidos vasculares en pastos y leguminosas como: el xilema y esclerénquima más abundantes en los tallos. Estos tejidos son bastante difíciles de degradar por la microbiota ruminal (Grabber el al. 1992) e incluyen una gran proporción de la fracción no digestible de un forraje.
Otros tejidos, como diferentes tipos de parénquima y tejido mesófilo no son lignificados y contienen una gran proporción de metabolitos altamente degradables. Otros son medianamente degradables, tales como el phloema y collenchyma (Wilson, J R et al, 1989, 1993).
La digestibilidad de un forraje representa las contribuciones de los varios tejidos en ese forraje al contenido de la materia seca en ese forraje. La mayor proporción que un forraje tenga en tejidos lignificados y otros carbohidratos estructurales daría como resultado que ese forraje tenga una menor digestibilidad. Son las diferencias genéticas entre especies forrajeras, las que dictan la distribución de lignina y otros componentes en la pared celular. Pastos del tipo tropical tienen una mayor concentración de lignina en sus tejidos que pastos de clima templado y por lo tanto, pastos de clima templado son más digestibles.
Morfología.-
La lignificación se incrementa conforme la planta madura. Esto es una función debida a cambios en su morfología y en menor grado al envejecimiento que experimenta. En los pastos el desarrollo morfológico se divide en 4 estados de crecimiento: a) vegetativo, b) alongamiento, c) reproductivo y e) maduración de las semillas.
Durante el periodo vegetativo de crecimiento, los tallos permanecen cerca de la corona y la porción más arriba de la corona consiste principalmente de hojas. Durante el alongamiento los entre nudos del tallo se alargan e incrementan en proporción al número de hojas ya formado.
Inflorescencia es el tercer periodo de desarrollo y es cuando la fertilización ocurre y semillas se desarrollan.
En leguminosas y otras dicotiledóneas ocurren los mismos estados de desarrollo. Durante el estado de desarrollo vegetativo los tallos crecen al mismo tiempo que las hojas y acumulación de tejido del tipo xilema se acumula durante este continuo desarrollo. Conforme plantas maduran el radio entre hojas a tallos disminuye. Los tallos contienen una mayor acumulación de lignina en sus tejidos y una menor cantidad de tejidos metabólicamente activos como los existentes en las hojas.
Referencias:
Akin, D.E., and D. Burdick. 1975. Percentage of tissue types in tropical and temperate grass leaf blades and degradation of tissues by rumen microorganisms. Crop Sci15: 661-669.Akin, D.E., J R. Wilson, and W.R.
Akin, D.E. 1989. Histological and physical factors affecting digestibility at forages. Agron. J. 81:17–25.
Allen MS, Mertens DR. Evaluating constraints on fiber digestion by rumen microbes. J Nutr. 1988;118:261–270.
Bae, H. D., T. A. McAllister, L. J. Yanke, K.-J. Cheng and A. D. Muir. 1993. Effect of condensed tannins on endoglucanase activity and filter paper digestion by Fibrobacter succinogenes S85. Appl. Environ. Microbiol. 59:2132-2138.
Ben-Ghedalia, D., A. Halevi, and J. Miron. 1995. Digestibility by dairy cows of monosaccharide components in diets containing wheat or ryegrass silage.
Boudet, A. M. 1998. A new view of lignification. Trends in Plant Sci. 3:67–71.
Buscaglia, H.J., H.M. Van Es, L.D. Geohring, H.C.A.M. Vermeulen, G.W. Fick, and R.F. Lucey. 1994. Alfalfa yield and quality are affected by soil hydrologic conditions. Agron. J. 86:535–542
Buxton, D.R. and J.R. Russell. 1988. Lignin constituents and cell-wall digestibility of grass and legume stems. Crop Sci. 28:553–558
Buxton, D.R. 1989. In vitro digestion kinetics of temperate perennial forage legume and grass stems. Crop Sci. 29:213–219.
Buxton, D.R. and M.R. Brasche. 1991. Digestibility of structural carbohydrates in coolseason grass and legume forages. Crop Sci. 31:1338–1345.
Buxton, D.R. and M.D. Casler. 1993. Environmental and genetic effects on cell wall composition and digestibility. p. 685–714.
Casler, M.D. and H.G. Jung. 1999. Selection and evaluation of smooth bromegrass clones with divergent lignin or etherified ferulic acid concentrations. Crop Sci. 39:1866–1873.
Cherney, J.H., D.J.R. Cherney, D.E. Akin, and J.D. Axtell. 1991. Potential of brown-midrib, low-lignin mutants for improving forage quality. Advances in Agron. 46:157–198.
Chesson, A., C. S. Stewart and R. J. Wallace. 1982. Influence of plant phenolics acids on growth and cellulolytic activity of rumen bacteria. Appl. Environ. Microbiol. 44:597-603
Chesson, A. 1993. Mechanistic models of forage cell wall degradation. In p. 347-376. H. G. Jung, et al. (eds.). Forage Cell Wall Structure and Digestibility. ASA-CSSA-SSSA, Madison, Wisc
Dehority, B. A. 1993. Microbial ecology of cell wall fermentation. In: Forage Cell Wall Structure and Digestibility
Engels, F.M., and H.G. Jung. 1998. Alfalfa stem tissues: cell-wall development and lignification. Ann. Bot. 82:561–568
Fahey, G.C., Jr., L.D. Bourquin, E.C. Titgemeyer, and D.G. Atwell. 1993. p. 715- 766. In H. G. Jung, D. R. Buxton, R. D.
Hatfield, and J. Ralph (ed.) Forage cell wall structure and digestibility. ASA-CSSA-SSSA, Madison, Wisc
Grabber, J.H., J. Ralph, and R.D. Hatfield. 1998. Ferulate cross-links limit the enzymatic degradation of synthetically lignified primary walls of maize. J. Agr.. Food Chem. 46:2609–2614.
Grabber, J.H., G.A. Jung, S.M. Abrams, and D.B. Howard. 1992. Digestion kinetics of parenchyma and sclerenchyma cell walls isolated from orchardgrass and switchgrass. Crop Sci. 32:806–810.
Halim, R.A., D.R. Buxton, M.J. Hattendorf, and R.E. Carlson. 1989. Water-stress effects on alfalfa forage quality after adjustment for maturity differences. Agron. J. 81:189–194.
Heinrichs AJ Kononoff PJ. Evaluating particle size of forages and TMRs using the new Penn state forage particle separator. Tech. Bull. DAS 96–20. Pennsylvania State University, College Agric. Sci., Cooperative Ext., University Park, PA; 2002.
Hungate, R. E. 1966. The Rumen and Its Microbes. Academic Press, New York
D. O. Krause,* T. G. Nagaraja,† A. D. G. Wright,‡ and T. R. 2014. Callaway§Rumen microbiology: Leading the way in microbial ecology1,2
Jung, H.G., and M.S. Allen. 1995. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J Anim Sci 73: 2774-2790.
Jung, H.G. and D.A. Deetz. 1993. Cell wall lignification and degradability. p. 315-346. In H. G. Jung, et al. (eds.). Forage Cell Wall Structure and Digestibility. ASA-CSSA-SSSA, Madison, Wisc.
Jung, H.G. and K.P. Vogel. 1992. Lignification of switchgrass (Panicum virgatum) and big bluestem (Andropogon gerardii) plant parts during maturation and its effect on fiber degradability. J. Sci. Food Agr. 59:169–176.
Jung, H.-J. G. and T. Sahlu. 1986. Depression of cellulose digestion by esterified cinnamic acids. J. Sci. Food Agric. 37:659-665
Klopfenstein, T. 1978. Chemical treatment of crop residues. J. Anim. Sci. 46:841–848.
Lennox, J. G., ed. 2001. Aristotle: Animalium partibus. Oxford Univ.Press, New York.
McCormick, M. E., J. F. Beatty, and J. M. Gillespie. 2002. Ryegrass bale silage research and management practices. Louisiana Agricultural Experiment Station Research Summary No. 144.
Mertens, D.R. 1994. Regulation of forage intake. p. 450-493. In G. C. Fahey, Jr. (ed.) Forage Quality, Evaluation, and Utilization. ASA, CSSA, SSSA, Madison, Wisc.
Miller, D.A. and H.F. Reetz, Jr. 1995. Forage fertilizer. p. 71–87. In R. F. Barnes, D. A. Miller, and C. J. Nelson (eds.) Forages: An Introduction to Grassland Agriculture, 5th ed, Vol. I. Iowa State University Press, Ames, Iowa.
Moore, K.J., and H.G. Jung. 2001. Lignin and fiber digestion. J Range Mgmt 54: 420- 430.
Morrison, T.A., H.G. Jung, D.R. Buxton, R.D. Hatfield. 1998. Cell-wall composition of maize internodes of varying maturity. Crop Sci. 38:455-460
Ramalho, J.M.C., 1991. Treatment of straws. Options Méditerranéennes 16: 55-60. treatment of low quality forages. Nebraska Cooperative Extension EC 89-265.
Stone, B.A. 1994. Prospects for improving the nutritive value of temperate, perennial pasture grasses. New Zealand J Agric Res 37: 349-363.
Sundstizil, F. 1988. Straw and other fibrous by-products. Livestock Prod Sci 19: 137-1 58
Van Soest, P.J. 1964. Symposium on nutrition and forage and pastures: New chemical procedures for evaluating forages. J. Anim. Sci. 23:838–84
Varga, G, Ihler V. Penn State. Carbohydrate nutrition for lactating dairy cattle
Vogel, K.P. and D.A. Sleper. 1994. Alteration of plants via genetics and plant breeding. p. 891–921. In: G. C. Fahey, Jr. (ed.) Forage Quality, Evaluation, and Utilization. ASA, CSSA, SSSA, Madison, Wisc
Waldo DR, Smith LW, Cox EL. Model of cellulose disappearance from the rumen. J Dairy Sci. 1972;55:125–129.
Wilson, J.R., and J. B. Hacke. 1987. Comparative digestibility and anatomy of some sympatric C3 and C4 arid zone grasses. Aust J Agric Res 38: 287-295.
Wilson, J.R.. 1993. Organization of forage plant tissues. p. 1–32. In: H. G. Jung, D. R. Buxton, R. D. Hatfield, and J. Ralph (ed.) Forage cell wall structure and digestibility. ASA-CSSASSSA, Madison, Wisc.
Wilson, J.R. and P.W. Hattersley. 1989. Anatomical characteristics and digestibility of leaves of Panicum and other grass genera with C3 and different types of C4 photosynthetic pathway. Aust. J. Agr. Res. 40:125–136.
Windham. 1983. Site and rate of tissue digestion in leaves of C4 and C3/C4, intermediate Panicum species. Crop Sci 23: 147-55.
Yang WZ, Beauchemin KA. Altering physically effective fiber intake through forage proportion and particle length: Chewing and ruminal pH. J Dairy Sci. 2007;90:2826–2838