Importancia de la diferencia catión-anión en la alimentación
Publicado: 29 de noviembre de 2006
Por: Andrés L. Martínez Marín, Veterinario
INTRODUCCIÓN.
Los avances en la alimentación de rumiantes han permitido una mejor estimación de las necesidades y aportes de los nutrientes: proteína, carbohidratos, grasa, vitaminas y minerales. El esfuerzo investigador para dividir los nutrientes “mayores” en subgrupos ha hecho posible que la formulación de las raciones sea más precisa por un mejor conocimiento de las interrelaciones entre los diferentes subgrupos.
En el caso de los minerales, la variedad de componentes que integran el grupo y la diversidad de funciones biológicas en las que participan ha dificultado la estimación precisa de los aportes necesarios de cada uno de ellos, y sobre todo, el conocimiento de los efectos que sus interrelaciones tienen en el organismo animal. Hoy en día aún se agrupan los minerales en dos grandes grupos: macro y microminerales; esta división está basada en la concentración de los minerales en el organismo animal y carece de significado biológico, no aportando ninguna información válida para la formulación de raciones.
El concepto de la diferencia o balance catión-anión de la dieta (DCAD) ha supuesto un avance en el conocimiento de los efectos fisiológicos que las interrelaciones entre los minerales tienen en el organismo animal y ha ayudado a la mejora de la productividad y a la prevención de trastornos metabólicos de elevado costo económico en el ganado vacuno lechero.
FUNDAMENTOS DE LA DIFERENCIA CATIÓN-ANIÓN.
Las interrelaciones entre los minerales sodio (Na) y potasio (K) –cationes- y el cloro (Cl) –anión-, pueden expresarse como su balance en el organismo animal. La premisa básica es que los tejidos vivos son eléctricamente neutros y que el equilibrio ácido-básico está determinado por la ingestión y excreción de estos iones que se denominan “iones fijos” ya que no pueden ser metabolizados a compuestos inferiores.
La DCAD de los alimentos o de las raciones se calcula en miliequivalentes por kilo o por 100 gramos de materia seca según la ecuación general (Na + K) – (Cl). Los iones están expresados en miliequivalentes. La conversión de porcentaje a miliequivalentes es: (%Na/0.023 + %K/0.039) - (%Cl/0.0355), y el resultado vendría dado en miliequivalentes por cada 100 gramos de materia seca. El ión sulfato, aunque no es un ión fijo, puede acidificar directamente los fluidos orgánicos y alterar el equilibrio ácido-básico si se incorpora en altas proporciones en la ración. Por ello, el ión sulfato debe incorporarse en la ecuación anterior si se añade deliberadamente a la ración (p. ej. en raciones de vacas secas).
La elección de estos minerales particulares (Na, Cl, K) para el cálculo de la DCAD, es debida a su participación en el balance osmótico del organismo, equilibrio ácido-básico e integridad de los mecanismos de transporte de las membranas celulares. Se entiende que el cálculo es válido cuando los minerales se encuentran en la ración en valores normales, ni deficientes ni tóxicos. Para la manipulación de la DCAD se utilizan sales minerales: bicarbonato sódico y potásico, fosfato sódico, sulfatos y cloruros de calcio, de magnesio o de amonio, etc.
Un concepto que origina confusión es que los aniones forman residuos ácidos (es decir, son acidogénicos), y que los cationes originan residuos alcalinos (es decir, son alcalogénicos). Sin embargo, este concepto es erróneo: ej. el ión fosfato (HPO4=) y el ión amonio (NH4+) actúan como donantes de protones aunque uno es un anión y otro un catión; igualmente no hay reacciones en las que el sodio o el potasio formen álcalis, no obstante estos iones, al igual que el cloro y el sulfato, afectan indirectamente a la concentración de iones hidrógeno (H+) en el organismo a través de los sistemas tampón orgánicos, la función renal y la respiración celular.
Por tanto, la DCAD, tal como es descrita por la ecuación (Na + K) – (Cl), no determina las propiedades acidogénicas o alcalogénicas de una ración o un alimento pero se puede utilizar para valorar el efecto que la absorción y el metabolismo de los iones considerados tienen sobre los sistemas tampón, la función renal y la respiración celular.
INFLUENCIA DEL EQUILIBRIO IÓNICO EN LOS PROCESOS METABOLICOS.
La influencia de los iones sobre los procesos metabólicos es compleja pero su conocimiento es necesario para comprender el efecto que la alteración de la DCAD tiene sobre los resultados productivos y estado sanitario de los animales. A continuación se resumen los cinco procesos metabólicos en que los iones influyen directa o indirectamente.
A) La bomba de sodio-potasio.
Este mecanismo celular mantiene constantemente altos niveles de potasio y bajos niveles de sodio dentro de la célula, el proceso requiere energía en forma de adenosín-trifosfato (ATP). De hecho, se estima que este proceso supone el 40% del gasto energético celular. La bomba de sodio-potasio funciona constante e independientemente de otros procesos metabólicos. El transporte de iones a través de la membrana celular se hace en conjunción con el transporte de glucosa al interior de la célula (figura I). Considerando que la glucosa es la principal fuente de energía celular, es obvio que la ralentización de la bomba impedirá que la célula funcione a su máxima capacidad. Esto es particularmente importante en las células de la glándula mamaria donde se requieren elevadas cantidades de glucosa para la síntesis de lactosa. Es evidente que la presencia de un catión en exceso sobre el otro provocará una reducción del mecanismo de bombeo, lo que ahorrará energía a la célula pero simultáneamente obtendrá menor cantidad de glucosa, o aumentará la velocidad de bombeo por encima del nivel óptimo con el consiguiente derroche de energía.
Figura I / ESQUEMA DE LA BOMBA DE SODIO-POTASIO
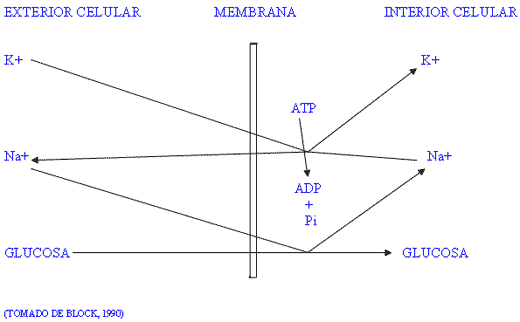
Figura I / ESQUEMA DE LA BOMBA DE SODIO-POTASIO
B) La excreción renal de sodio y potasio.
En el riñón se conserva potasio a expensas de la eliminación de sodio. El exceso de un catión frente al otro puede provocar deficiencia inducida. Por ejemplo, si el potasio se encuentra en exceso, el riñón eliminará sodio creando una deficiencia a pesar de que el sodio se encuentre en la ración en cantidades adecuadas. En esta circunstancia, un déficit de cloro agravará aún más el problema ya que se requiere cloro para la eliminación renal de potasio en exceso.
C) El equilibrio ácido-básico de la sangre.
En el intestino delgado se absorbe cloro, cuando está en exceso sobre el sodio, por intercambio con el ión bicarbonato (HCO3-) para mantener la neutralidad eléctrica. Si la presencia de sodio es insuficiente para permitir la absorción de cloro y sodio en cantidad eléctricamente neutra, ocurrirá un excesivo paso de bicarbonato desde la sangre a intestino para compensar la absorción de cloro, que creará un estado de acidosis metabólica. Alternativamente, existe la posibilidad de intercambiar el sodio ingerido por el ión hidrógeno (H+) de la sangre cuando el sodio está en exceso sobre el cloro. Esta situación puede producir alcalosis metabólica. De hecho, el efecto alcalogénico de un exceso de sodio puede afectar negativamente a los animales si no está equilibrado con el cloro, esta situación es corriente cuando se añade bicarbonato sódico (NaHCO3) a las raciones de vacas lecheras sin alterar el aporte de sal (ClNa) para compensar el aporte extra de sodio.
D) La función renal.
Figura II / FUNCIÓN RENAL
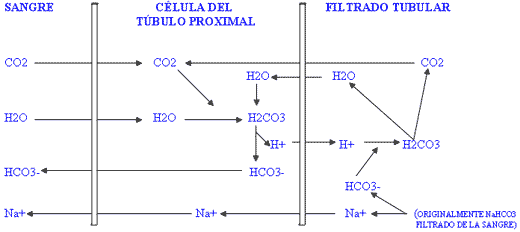
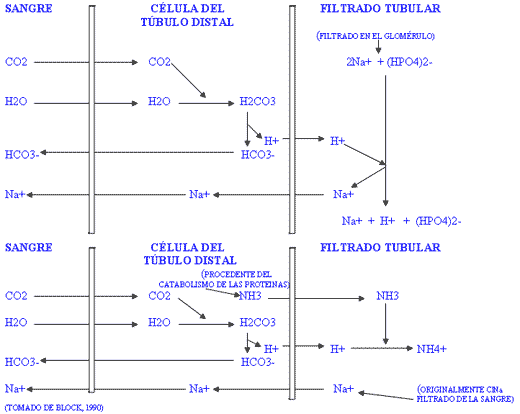
La movilización de ión hidrógeno en los túbulos proximales del riñón así como su excreción al filtrado glomerular y la formación de amoniaco en los túbulos distales, dependen de la reabsorción de sodio para compensar eléctricamente la reabsorción de bicarbonato desde las células tubulares a la sangre. Cuando existe acidosis leve en los fluidos extracelulares, los riñones conservan bicarbonato por reabsorción, lo contrario es cierto en caso de alcalosis. Si el cloro se encuentra en exceso en el filtrado glomerular, ocurrirá intercambio del mismo con el bicarbonato intracelular para la absorción de Cl-Na+, en consecuencia se reduce el paso de bicarbonato a la sangre. Por esto, un exceso de cloro impedirá que el organismo pueda reabsorber bicarbonato para contrarrestar una situación de acidosis leve.
La movilización de ión hidrógeno en los túbulos proximales del riñón así como su excreción al filtrado glomerular y la formación de amoniaco en los túbulos distales, dependen de la reabsorción de sodio para compensar eléctricamente la reabsorción de bicarbonato desde las células tubulares a la sangre. Cuando existe acidosis leve en los fluidos extracelulares, los riñones conservan bicarbonato por reabsorción, lo contrario es cierto en caso de alcalosis. Si el cloro se encuentra en exceso en el filtrado glomerular, ocurrirá intercambio del mismo con el bicarbonato intracelular para la absorción de Cl-Na+, en consecuencia se reduce el paso de bicarbonato a la sangre. Por esto, un exceso de cloro impedirá que el organismo pueda reabsorber bicarbonato para contrarrestar una situación de acidosis leve.
E) La respiración celular.
Figura III / CAMBIO MINERAL DE LOS ERITROCITOS DURANTE LA RESPIRACIÓN.
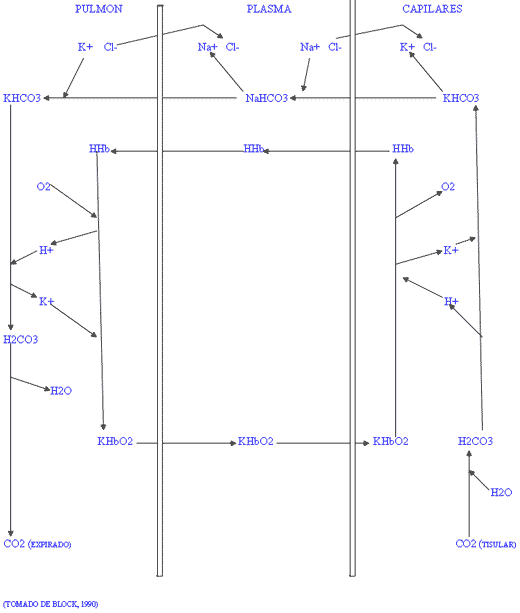
El principal tampón de la sangre es la sal potásica de oxihemoglobina (KHbO2) presente en el interior de los eritrocitos. El dióxido de carbono producido en el metabolismo tisular reacciona con agua para formar ácido carbónico (H2CO3) dentro de los eritrocitos. Parte del ácido carbónico pasa al plasma y el resto reacciona con la sal potásica de oxihemoglobina para formar bicarbonato con la liberación de oxígeno y potasio. El bicarbonato es intercambiado por cloro presente en el plasma sanguíneo, formándose bicarbonato sódico en el plasma y cloruro potásico en el interior de los eritrocitos, para mantener la neutralidad. La reacción es reversible en el pulmón donde el cloro retorna a la sangre liberando al potasio que forma nuevamente sal potásica de oxihemoglobina, el cloro neutraliza al sodio sanguíneo liberado cuando el bicarbonato reentra a los eritrocitos para la eliminación del dióxido de carbono. El desequilibrio de los iones puede originar alcalosis o acidosis por insuficiente intercambio de bicarbonato y ácido a través de la membrana de los eritrocitos.
RESPUESTA DE LAS VACAS LACTANTES A LA MANIPULACIÓN DE LA DCAD.
Con todo lo anteriormente expuesto es aparente que la mayor influencia de la DCAD es en la regulación del equilibrio ácido-básico. Este efecto no puede ser medido con el pH de la sangre porque éste es la suma de las reacciones totales del organismo y está altamente tamponado. La modificación del estado ácido-básico activa los mecanismos compensatorios orgánicos favoreciendo cambios fisiológicos que nos pueden interesar desde el punto de vista productivo.
La influencia de la manipulación de la DCAD en vacas lactantes es diferente según exista o no una situación de estrés por calor.
1) Efecto de la DCAD cuando no hay estrés por calor.
La manipulación de la DCAD afecta diversos parámetros productivos y fisiológicos de las vacas lactantes. El aumento de la DCAD por manipulación del contenido en sodio, potasio o cloro (cuadro I) aumenta el consumo de materia seca y la producción de leche. La producción de grasa no es modificada. La producción de proteína aumenta ligeramente. El pH ruminal aumenta linealmente sin que se altere el patrón de fermentación. El pH de la sangre tiende a aumentar debido a la pérdida de protones (H+) a nivel intestinal para compensar la mayor absorción de sodio y potasio. El pH urinario aumenta de forma lineal primero y luego de forma cuadrática. Desde el punto de vista fisiológico, el aumento del pH plasmático más allá de cierto punto conlleva el aumento del pH urinario ya que, para el mantenimiento de la homeostasis, el organismo compensa la pérdida de ácido en la sangre y la alteración de la relación bicarbonato a dióxido de carbono, aumentando la excreción renal de bicarbonato. Las concentraciones de sodio y potasio en la sangre no son significativamente afectadas por la DCAD, este fenómeno asociado a una mayor presencia de estos cationes en la orina y a una menor concentración de cloro en la sangre sugiere que ocurre una mayor excreción de cationes en la orina que se acompaña de la pérdida de cloro para mantener la neutralidad eléctrica a nivel renal. El hecho de que la respuesta sea igual, independientemente del ión manipulado, sugiere que la respuesta observada es achacable enteramente a la diferente DCAD de las raciones. Parece ser que la máxima ingesta de materia seca, producción de leche y producción de leche corregida al 4% de grasa se obtiene con una DCAD de +38 meq/100 gramos de materia seca, no obstante, la magnitud de la respuesta es pequeña entre +25 y +50 meq/100 gramos de materia seca (figura IV).
Cuadro 1 / EFECTOS DE LA DCAD SOBRE DIVERSOS PARAMETROS EN VACAS LACTANTES
Cuadro 1 / EFECTOS DE LA DCAD SOBRE DIVERSOS PARAMETROS EN VACAS LACTANTES
DCAD = (Na + K) - Cl, meq/100 gr ms | ||||||
-10 | 0 | 10 | 20 | |||
IMS KG/DIA | 16.8 a | 18 b | 18.3 b | 18.6 b | ||
LECHE KG/DIA | 18.5 a | 19.5 a | 19.6 ab | 20.1 b | ||
GRASA KG/DIA | 0.71 | 0.71 | 0.72 | 0.76 | ||
PROTEINA KG/DIA | 0.6 a | 0.66 b | 0.65 ab | 0.65 ab | ||
pH SANGRE | 7.34 a | 7.38 a | 7.42 b | 7.43 b | ||
HCO3 SANGUINEO meq/l | 19.3 a | 21.1 b | 22.2 c | 23.2 c | ||
PRESIÓN DE CO2 mmHg | 34.4 a | 37.1 b | 35.5 ab | 36.3 ab | ||
pH ORINA | 6.1 a | 6.1 a | 6.4 a | 7.6 b | ||
ACETATO RUMINAL % | 61 | 64 | 63.5 | 63.2 | ||
PROPIONATO RUMINAL % | 24.6 | 21.2 | 22 | 22.3 | ||
BUTIRATO RUMINAL % | 11.5 | 11.9 | 11.4 | 11.1 | ||
ISOVALERATO RUMINAL % | 1.5 a | 1.7 a | 1.7 a | 2.0 b | ||
A.G.V. TOTALES mmol/l | 104.9 b | 98.3 ab | 91.5 a | 94.8 ab | ||
pH RUMINAL | 6.45 a | 6.63 ab | 6.73 b | 6.76 b | ||
(LOS VALORES CON DIFERENTE LETRA INDICAN UNA DIFERENCIA SIGNIFICATIVA, p<0.05) | ||||||
(DE TUCKER ET AL. 1988) |
Figura IVa / EFECTO DE LA DCAD SOBRE EL CONSUMO
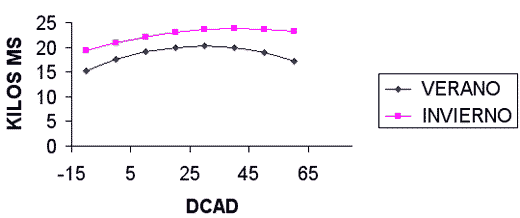
(SEGUN SANCHEZ ET AL, 1994)
Figura IVb / EFECTO DE LA DCAD SOBRE LA PRODUCCION DE LECHE
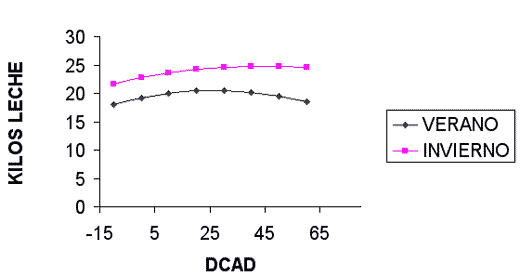
(SEGUN SANCHEZ ET AL, 1994)
(SEGUN SANCHEZ ET AL, 1994)
Figura IVb / EFECTO DE LA DCAD SOBRE LA PRODUCCION DE LECHE
(SEGUN SANCHEZ ET AL, 1994)
Debido a la elevada tasa metabólica al comienzo de la lactación, el organismo de la vaca tiende a estar acidótico, por ello el mantenimiento de una DCAD elevada (alcalogénica) en la ración determinará mejores resultados productivos en este período. Al avanzar los días en leche, la DCAD de la ración es menos importante porque el estado ácido-básico del organismo tiende hacia la alcalinidad (aumenta el pH, el bicarbonato y la presión parcial de dióxido de carbono en la sangre).
b) Efectos de la manipulación de la DCAD durante el estrés por calor.
Durante el estrés por calor ocurren cambios fisiológicos en el organismo cuya finalidad es el mantenimiento de la homeostasis. El jadeo, necesario para aumentar la pérdida de calor por una mayor evaporación desde los conductos respiratorios, altera la ventilación alveolar afectando a la presión parcial y a la concentración de dióxido de carbono, lo que modifica el pH de la sangre. El dióxido de carbono es eliminado más rápidamente de lo que se produce. El descenso de la presión parcial de dióxido de carbono crea un déficit de ácido carbónico en la sangre con el consiguiente aumento del pH sanguíneo. El organismo trata de compensar el mayor pH de la sangre aumentando la excreción renal de bicarbonato y reduciendo la excreción de ácido, lo que origina un incremento del pH urinario. La consecuencia final es una relación bicarbonato/presión parcial de dióxido de carbono y ácido carbónico/bicarbonato que permite mantener un pH sanguíneo normal. La alteración de la concentración de ácido carbónico y bicarbonato para compensar la alcalosis respiratoria origina acidosis metabólica por sobrecompensación. En definitiva, durante la parte cálida del día las vacas se encuentran alcalóticas (mayor pH en sangre y en orina), por el contrario, durante la parte más fresca del día, el pH sanguíneo y urinario son menores de lo normal en vacas no sometidas a estrés por calor.
La combinación de los efectos del estrés por calor en el equilibrio ácido-básico y la alteración del patrón de comidas en estas circunstancias puede tener un efecto profundo sobre los resultados productivos. El mayor consumo de alimento durante las horas más frescas del día ocasionará un aumento de la producción de ácidos en rumen justamente cuando está ocurriendo acidosis metabólica compensatoria, y la capacidad tampón de la saliva está disminuida. En estas circunstancias, deben modificarse las prácticas de alimentación.
Vistos los cambios fisiológicos que ocurren durante el estrés por calor, vamos a repasar cual es el papel de la DCAD en dichas circunstancias. La necesidad a nivel renal de eliminar bicarbonato y conservar iones hidrógeno para reducir el pH de la sangre, incrementa enormemente la demanda de cationes, ya que la excreción de bicarbonato requiere la presencia de un catión para mantener la neutralidad eléctrica. El sodio es más utilizado que el potasio. Durante el estrés por calor la concentración de potasio en la orina y en la sangre disminuye, lo que refleja la utilización del mismo para otras funciones como p. ej. la producción de sudor. La acidosis metabólica compensatoria requiere una menor presencia de cloro para favorecer la reabsorción de bicarbonato. El incremento en la demanda de sodio para la excreción renal, de potasio para la sudoración y la menor necesidad de cloro, son consistentes con la necesidad de una DCAD equilibrada, como lo demuestra el efecto cuadrático marcado de los tres iones sobre el consumo y la producción de leche durante el verano. Sin embargo, el potencial alcalogénico de una DCAD elevada determina un efecto menor del incremento de la misma sobre el consumo y la producción de leche en verano que en invierno.
Vistos los cambios fisiológicos que ocurren durante el estrés por calor, vamos a repasar cual es el papel de la DCAD en dichas circunstancias. La necesidad a nivel renal de eliminar bicarbonato y conservar iones hidrógeno para reducir el pH de la sangre, incrementa enormemente la demanda de cationes, ya que la excreción de bicarbonato requiere la presencia de un catión para mantener la neutralidad eléctrica. El sodio es más utilizado que el potasio. Durante el estrés por calor la concentración de potasio en la orina y en la sangre disminuye, lo que refleja la utilización del mismo para otras funciones como p. ej. la producción de sudor. La acidosis metabólica compensatoria requiere una menor presencia de cloro para favorecer la reabsorción de bicarbonato. El incremento en la demanda de sodio para la excreción renal, de potasio para la sudoración y la menor necesidad de cloro, son consistentes con la necesidad de una DCAD equilibrada, como lo demuestra el efecto cuadrático marcado de los tres iones sobre el consumo y la producción de leche durante el verano. Sin embargo, el potencial alcalogénico de una DCAD elevada determina un efecto menor del incremento de la misma sobre el consumo y la producción de leche en verano que en invierno.
Por último señalar que la utilización de bicarbonato sódico además de alterar la DCAD tiene un efecto específico sobre la concentración de grasa en la leche durante el estrés por calor o cuando las raciones son pobres en forraje largo. En ambos casos, el efecto es debido a la actividad tampón del bicarbonato en rumen que compensa la menor entrada de tampones con la saliva, y en consecuencia el menor pH ruminal, que ocurre en ambas situaciones.
INFLUENCIA DE LA DCAD EN LAS VACAS DURANTE EL PERIODO DE SECADO.
Los trastornos asociados con la hipocalcemia clínica o subclínica (“fiebre de la leche”, retención placentaria, cetosis, desplazamiento precoz de abomaso) tienen un elevado coste económico debido a los gastos veterinarios, producción disminuida y posible pérdida del animal. La prevención de la hipocalcemia se ha orientado en varios sentidos: reducción de los aportes de calcio y fósforo en las raciones de secado, estimulación de la absorción y utilización del calcio mediante tratamientos con vitamina D, y manipulación de la DCAD. En la presente revisión nos vamos a referir únicamente a la alteración de la DCAD como mecanismo para la prevención de la hipocalcemia en vacas.
Hoy día se considera más grave un excesivo aporte de potasio que de calcio como desencadenante de la hipocalcemia. El efecto de un exceso de potasio (de una DCAD elevada) es, como hemos visto, la creación de un estado alcalótico en el organismo. Esta alcalosis es la responsable de los bajos niveles de calcio en la sangre en el momento del parto. Se cree que los receptores celulares para paratohormona (en riñón y hueso) son temporalmente refractarios a su estimulación cuando el pH sanguíneo es elevado. Esta situación compromete la homeostasis del calcio. Cuando la DCAD es igual o inferior a cero (predominio de los aniones sobre los cationes, “ración aniónica”), se produce una acidosis metabólica leve (retención de iones hidrógeno) para compensar la absorción de cloro (y azufre). La acidosis metabólica favorece la acción de la paratohormona. Para mantener el pH de la sangre dentro de límites normales y aumentar el calcio en la sangre (demanda de formación de calostro y leche) se activan mecanismos compensatorios fisiológicos paralelos. La secuencia de eventos (figura V) se mantiene en tanto el animal siga consumiendo una ración rica en aniones.
Figura V / EVENTOS FISIOLOGICOS DURANTE EL PERIPARTO Y EFECTO DE LA DCAD NEGATIVA.
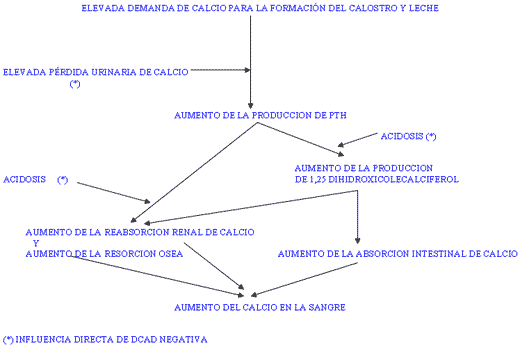
Los niveles que se consideran óptimos para la DCAD de vacas secas son de –5 a –20 meq/100 gramos de materia seca. La medida más fácilmente determinable de la DCAD conseguida en la ración es el pH urinario, existiendo la relación: pH = 6.304 + (0.05* DCAD), donde DCAD está expresado en meq/100 gr ms. La DCAD es una medida adecuada para la valoración de raciones de secado que permita prevenir la hipocalcemia (cuadro II).
Cuadro II / RESULTADOS DE LA UTILIZACIÓN DE DIFERENTE DCAD EN VACAS DURANTE EL PREPARTO
Figura V / EVENTOS FISIOLOGICOS DURANTE EL PERIPARTO Y EFECTO DE LA DCAD NEGATIVA.
Los niveles que se consideran óptimos para la DCAD de vacas secas son de –5 a –20 meq/100 gramos de materia seca. La medida más fácilmente determinable de la DCAD conseguida en la ración es el pH urinario, existiendo la relación: pH = 6.304 + (0.05* DCAD), donde DCAD está expresado en meq/100 gr ms. La DCAD es una medida adecuada para la valoración de raciones de secado que permita prevenir la hipocalcemia (cuadro II).
Cuadro II / RESULTADOS DE LA UTILIZACIÓN DE DIFERENTE DCAD EN VACAS DURANTE EL PREPARTO
DCAD = (Na + K) - (Cl +S), meq/100 gr | |||||||
"-25 meq/100 gr" | "+5 meq/100 gr" | ||||||
MEDIDA | |||||||
HIPOCALCEMIA CLINICA % | |||||||
< 3 LACTACIONES | 0 | 0 | |||||
> 3 LACTACIONES | 5 | 12 | |||||
TODAS | 4 | 9 | |||||
HIPOCALCEMIA SUBCLÍNICA %(*) | |||||||
< 3 LACTACIONES | 2 | 16 | |||||
> 3 LACTACIONES | 28 | 66 | |||||
TODAS | 19 | 50 | |||||
TODAS LAS VACAS | |||||||
Ca IÓNICO EN SUERO mg/dl | 4.31 | 3.8 | |||||
CALCIO EN SUERO mg/dl | 7.94 | 7.1 | |||||
FOSFORO EN SUERO mg/dl | 4.44 | 3.64 | |||||
PRODUCCION DE LECHE, 305 DIAS | 9300 | 9000 | |||||
PORCENTAJES DE CONCEPCIONES | |||||||
100 DIAS | 35 | 28 | |||||
150 DIAS | 55 | 42 | |||||
200 DIAS | 71 | 54 | |||||
250 DIAS | 77 | 66 | |||||
SERVICIOS POR CONCEPCIÓN | 3 | 3.4 | |||||
DIAS AL PRIMER CELO | 68 | 72 | |||||
DIAS ABIERTOS | 124 | 138 | |||||
(*) CONCENTRACIONES DE Ca2+ INFERIORES A 4 mg/dl SE CONSIDERAN INDICADORAS DE HIPOCALCEMIA SUBCLÍNICA | |||||||
(DE BEEDE ET AL 1992) |
La manipulación de la DCAD podría tener efectos a varios niveles:
- Función renal.
La DCAD negativa provoca un aumento de la eliminación de calcio en la orina. Este efecto podría ser debido a:
-- la reducción marcada del pH urinario por aumento de la excreción neta de ácidos que inhibe los mecanismos de reabsorción tubular de calcio
-- formación de complejos de sulfato y calcio en el filtrado glomerular que impiden la reabsorción de calcio, ya que el sulfato se reabsorbe en una tasa mínima
-- mayor disponibilidad de calcio en la sangre, ya que la DCAD negativa aumenta el nivel de calcio iónico en el plasma
-- la reducción marcada del pH urinario por aumento de la excreción neta de ácidos que inhibe los mecanismos de reabsorción tubular de calcio
-- formación de complejos de sulfato y calcio en el filtrado glomerular que impiden la reabsorción de calcio, ya que el sulfato se reabsorbe en una tasa mínima
-- mayor disponibilidad de calcio en la sangre, ya que la DCAD negativa aumenta el nivel de calcio iónico en el plasma
La pérdida de calcio en la orina estimula la producción de paratohormona (PTH) en la glándula paratiroides. Esta hormona estimula a su vez la producción renal de 1,25 dihidroxicolecalciferol (1,25(OH)2D3). La sensibilidad de las células renales productoras de 1,25 dihidroxicolecalciferol al estímulo de la paratohormona se ve favorecida por la acidosis metabólica moderada.
b) Resorción ósea.
El exceso de cloro presente en intestino cuando la DCAD es reducida, impide la reabsorción de bicarbonato en los riñones y aumenta su pérdida a nivel intestinal, lo que anula este mecanismo compensatorio de la acidosis metabólica provocada. En esta circunstancia, los huesos actúan como fuente de carbonato. En el proceso ocurre movilización pasiva de calcio y fósforo desde los huesos. Aparte de este mecanismo indirecto de estimulación de la movilización ósea, existe otro mecanismo específico, hormonal, que ocasiona resorción ósea. La calciuria debida a la DCAD negativa promueve la secreción de paratohormona y ésta estimula a su vez la producción de 1,25 dihidroxicolecalciferol. Ambas actúan sinérgicamente estimulando a los osteoclastos para activar la resorción del tejido óseo y liberar calcio a la sangre. Como la actividad de los osteoclastos es un proceso ácido-dependiente, una DCAD elevada traducida en un exceso de cationes (que causa deficiencia de iones hidrógeno) o en una falta de aniones (que produce un exceso de bicarbonato) alterará su funcionalidad limitando la resorción y el paso de calcio a la sangre. Si la DCAD es reducida, la acidosis metabólica consecutiva favorecerá la resorción mediada por hormonas y aumentará el calcio disponible en la sangre. Una vaca alimentada con forraje puede movilizar hasta 6-10 gr de calcio, esta cantidad aumenta 6-8 gr adicionales si la ración es aniónica. Por comparación, la formación del calostro requiere 2.3 gr/litro.
c) Aumento de la absorción de calcio en intestino.
Cuando la presencia de calcio en intestino es elevada, la acidificación directa de la ración aumenta la absorción pasiva de calcio, posiblemente por una mejor solubilización del mismo, pero no existe evidencia de que sales minerales neutras que aporten cantidades elevadas de aniones, reduzcan el pH en la luz intestinal. De hecho, lo más probable es lo contrario, el exceso de cloro prevendrá la salida de iones hidrógeno desde la sangre a la luz intestinal. Por otro lado, los animales pueden absorber calcio en intestino de forma activa cuando el aporte es inferior a las necesidades o muy superior a las mismas. En este caso, ocurre absorción activa mediada por la 1,25 dihidroxicolecalciferol. Esta hormona estimula la actividad de la Proteína Fijadora de Calcio y la ATP-asa-cálcica magnesio-dependiente presentes en el epitelio intestinal. La absorción de calcio intestinal en vacas consumiendo raciones con DCAD negativa ha sido variable, en unos casos ha ocurrido aumento de la absorción y en otros no se ha observado ninguna variación sobre el control. Posiblemente las discrepancias observadas sean debidas a las diferentes DCAD usadas y al nivel de calcio aportado.
CONCLUSIÓN.
CONCLUSIÓN.
La manipulación de la DCAD se constituye en una herramienta súmamente útil para favorecer la producción de las vacas lactantes o para prevenir trastornos metabólicos durante el periparto.
Durante la lactación, el efecto de un aumento de la DCAD es más aparente en invierno que en verano y más en vacas al comienzo de la lactación que al avanzar la misma. La DCAD óptima se sitúa en torno a 38 meq/100 gr de ms. Dentro de la DCAD conseguida se requiere que las relaciones sodio/potasio y sodio/cloro estén equilibradas.
La utilización de una DCAD negativa durante el período de secado permite mantener elevados niveles de calcio en la sangre durante el periparto evitando la hipocalcemia. El tiempo requerido de consumo de la ración aniónica es de al menos 10 días antes del parto y el efecto se mantiene durante aproximadamente 1 semana después del cambio de la ración. El uso de raciones aniónicas exige modificar los aportes de calcio y fósforo según la DCAD conseguida y vigilar los aportes de cloro y azufre para evitar el riesgo de toxicidad.
Artículo publicado en Mundo Ganadero nº 128. Diciembre de 2000.
© Mundo Ganadero. El mensual Mundo Ganadero lo edita Eumedia, S.A. en Madrid. C/ Claudio Coello, 16. 28001 MADRID.
Teléfono: (34) 91 426 44 30. Fax: (34) 91 575 32 97. e-mail: em@eumedia.es
Teléfono: (34) 91 426 44 30. Fax: (34) 91 575 32 97. e-mail: em@eumedia.es
Temas relacionados:
Autores:

Universidad de Córdoba - España
Recomendar
Comentar
Compartir

Universidad de Córdoba - España
20 de diciembre de 2006
Estimados compañeros:
Gracias por sus comentarios, le dejo unas aclaraciones que espero sean de su interés.
a) Respecto al ajuste de la diferencia catión-anión en vacas de ordeño: si las vacas se alimentan con raciones muy ricas en forrajes (sobre pastos o a pesebre), es de interés conocer la DCAD de dichos alimentos (seguramente se alcancen valores muy elevados ya que los forrajes suelen ser muy ricos en potasio). A partir de aquí decidiremos si conviene manipular la DCAD del concentrado que estemos aportando (básicamente incorporando bicarbonato sódico). Debe quedar claro que la repuesta positiva en consumo de materia seca y producción de leche alcanza un máximo en torno a 40 meq/100 gramos de materia seca, y que este valor se consigue rápidamente con raciones muy forrajeras. Una DCAD elevada más allá de dicho punto es contraproducente desde el punto de vista productivo.
b) Las vacas en periparto son susceptibles de padecer hipocalcemia clínica o subclínica si consumen raciones con forrajes muy ricos en potasio. En estas circunstancias sería útil el empleo de sales aniónicas para reducir la DCAD de la ración hasta valores de -7 a -15 meq/100 gramos de materia seca (ajustando claro está los aportes de calcio, fósforo y magnesio). Sin embargo, conseguir dicha reducción es en extremo difícil pues se parten de valores muy altos (en ocasiones pueden llegar a +100 meq/100 gramos de materia seca). Por otra parte, debido a la poca palatabilidad de las sales aniónicas, su incorporación en los concentrados a dosis elevadas puede provocar rechazo de los mismos con depresión del consumo total de energía, lo que complicaría aún más la situación (mi experiencia en este sentido es que la paridera puede acabar con resultados catastróficos). En definitiva, modificar la DCAD hasta valores favorables en las raciones de vacas en preparto requiere, si es posible, una selección previa de materias primas (principalmente forrajes) con bajo contenido en potasio para componer dichas raciones, de esta manera ajustaremos el aporte de las sales aniónicas al concentrado a la menor cantidad necesaria, para evitar rechazos en el consumo del mismo. Por cierto, el seguimiento de los resultados pueden hacerse fácilmente mediante la comprobación del pH de la orina (debe ser ligeramente ácido) con tiras reactivas.
c) Respecto a la manipulación de la DCAD en vacas nodrizas, no dispongo de información al respecto ni he tenido noticias de problemas relacionados con este tema, pero seguramente debido a la menor exportación de calcio en la leche (por la menor producción diaria), no sean necesarias medidas especiales de prevención de la hipocalcemia. En el caso de vacas con terneros debajo, aún es menos probable la necesidad de ajustar la DCAD ya que habitualmente estas vacas se mantienen sobre pastos, y como hemos dicho, los pastos y forrajes suelen tener diferencias catión-anión elevadas.
Para ampliar la información, les indico tres referencias fundamentales sobre este tema:
Effect of anionic salts in prepartum diets based on alfalfa. Joyce et al. J Dairy Sci 80:2866-2875
Effects of the addition of potassium or sodium but not calcium, to prepartum rations on milk fever in dairy cows. Goff et al. J Dairy Sci 80:176-186
Dietary cation-anion difference effects on performance and acid-base status of lactating dairy cows: a meta-analysis. Hu et al. J Dairy Sci 87:2222-2229
FELIZ NAVIDAD Y PAZ A LOS HOMBRES DE BUENA VOLUNTAD
Recomendar
Responder


19 de diciembre de 2006
BUEN DIA A TODOS ESPERANDO QUE TENGAN UNA FELIZ NAVIDAD Y PROSPERO AÑO NUEVO...QUISIERA FELICITAR EL ARTICULO Y ME GUSTARIA SABER COMO FUNCIONARIA EN GANADO DE CARNE SI SE TIENE ALGUN DATO SOBRE ESTO Y SOBRE LA FELICIDAD DE LOS ANIMALES SIMPLEMENTE SI TE DAN BUENA PRODUCCION ES QUE SE TIENE UN BUEN MANEJO BUENAS INSTALACIONES EN DONDE LOS ANIMALES ESTAN PLANAMENTE COMODOS Y ASI PUEDAN PRODUCIR LO QUE SE BUSCA. YA HAY METODOS QUE PERMITEN TENER MAYOR PRODUCCION MENOS ESTRES CON MUSICA ESO ESTA COMPROBADO EN VACAS LECHERAS Y A FIN DE CUANTAS TODOS TIENEN ALGUN FIN QUE PRODUCIR SI NO PUES DONDE ESTA LA CADENA ALIMENTICIA..PERO SI LO MEJOR ES TENER ANIMALES EN UN HABITAD DONDE ELLOS TENGAN MENOS ESTRES Y UNA LARGA VIDA QUE ES LO QUE BUSCAMOS TANTO LOS CONSUMIDORES COMO LOS PROVEDORES.
Recomendar
Responder

Phileo by Lesaffre
17 de diciembre de 2006
Doctor Martínez: Antes que nada, mis más sinceras felicitaciones por el artículo, muy interesante y completo. Al señor que pregunta por la felicidad y tranquilidad de las vacas, me permito decirle con todo respeto que mientras estemos trabajando por ellas desde lo nutricional, sanitario y de confort, con seguridad que se sentirán felices. En Colombia, especialmente en la zona norte de Antioquia, donde se concentra la mayor cantidad de hatos productores de leche con ganado especializado, se ha hablado mucho sobre la utilización de sales aniónicas durante el período de transición, pero, en la ganadería de trópico bajo, Córdoba, Sucre y Cesar, donde se produce el mayor volumen de leche bajo el sistema de Doble Propósito, el tema pocas veces se habla. Es por ello que me gustaría saber qué tanta influencia tiene la manupulación de las dietas en ganados tipo Cebú cruzados con ganados Holstein, Pardo entre otros, y bajo sistemas de pastoreo en Brachiarias y Dichiantium aristatum?? Bajo sistemas de pastoreo contínuo y alternado, y con niveles bajos de suplementación de alimentos balanceados?? Muchas gracias y FELIZ NAVIDAD a todos los colegas.
LAPL
Recomendar
Responder

22 de febrero de 2007
Agradezco a Carlos Meza por su solidaridad. Y aprovecho para agregar. Modernamente se impone cada vez más que una buena parte de los logros humanos provienen de la intuición. Pero la intuición no sigue caminos lógicos... aparentemente. Quizás recurra a los llamados caminos irracionales. De esta manera nos aproximamos más a la manera de pensar de los animales que deciden en función de formas situacionales (Ver a la autista Dra. Grandiner). El viejo fraude de que los animales no piensan, ni sienten, quedó en el pasado. El enorme impacto que tienen los perros de compañía sobre sus dueños a los que dispensan un amor casi includicable es una metáfora de lo que en general debería hacerse. ¡Y hay que ver la felicidad que logran esas personas haciendo felices a sus animales, y no viéndolos como las libras de carne que lograré para obtener mayores ganancias. Ojo, esto no es una cuestión religiosa. Ni es cosa de santurrones o cosa parecida. Es un asunto en el que va el porvenir de la humanidad. Y si no que esperemos lo que nos dirá el Cambio Climático. Allí los animales nos darán una lección, pero es posible que ya sea demasiado tarde. La clave de todo este pensamiento es que debemos abandonar las formas lógicas de pensamiento. Confiar más en la intuición. No existe la idea, por ejemplo: siempre existe un racimo de ideas. Kant y Hegel, están fuera de contexto. Toda la cuántica los mandó de paseo. Adiós a la Mecánica Clásica. Adios a Darwin. ¡Todas las especies son superiores, según el habitat en que viven! Es de esto que debemos alimentarnos para poder alimentar a los animales. ¿O es que todavía somos cartesianos?
Recomendar
Responder


21 de febrero de 2007
Diferencia o balance catión-anión en la dieta de bovinos
Dr. Martínez: Su artículo es muy importante ya que está enfocado a la fisiología celular y su influencia en el funcionamiento metabólico del aparato digestivo.
Es claro que desde que se suministra concentrado se han venido manipulando y controlando los procesos digestivos y alimenticios de los animales, buscando con ello el mayor beneficio en la productividad de éstos.
En este artículo resalta el control o manejo de la dieta catión-anión y su funcionamiento fisiológico en la digestión y su influencia en el proceso productivo. Nada nuevo está ocurriendo dentro del proceso de producción.
En cuanto a la felicidad de los animales, desde que el Hombre en su afán de conseguir dinero acostó de la felicidad de otros, ésta ha sido violada y vulnerada. Aquí me identifico con el paisano Tamara Gómez.
La felicidad: Es el estado de ánimo del que disfrutan los animales en su habitat natural. Corren, brincan, saltan y comen cuando quieren y lo que quieren, con cero estrés.
Que es diferente al bienestar u confort que es suplir o brindarles las necesidades físicas, sociales y etológicas claves de ellos, como manejo apropiado, salud, y algunas normas éticas que deben tener para con ellos, los dueños y criadores.
Esta es responsabilidad del veterinario, del dueño y de quién los tiene a su cuidado.
Aquí discrepo con el colega zootecnista LAPL.
Enrique C Meza A
Recomendar
Responder
20 de diciembre de 2006
Quiero agradecer a la Ingeniera María Rojas su referencia de la Dra. Grandin. He visitado esa maravillosa página. Me ha servido esa visita, además, para mejorar mis conocimientos sobre autismo. Es indudable que el manejo dispensado por la Dra. Grandín a los animales dista mucho del que comúnmente se les ofrece. Sin embargo, creo que esta temática, lo de la felicidad de los animales, que otros dan con el eufemismo de confort, no es para esta página a juzgar por el poco interés de los señores del estudio que no han dicho esta boca es mía. Están muy concentrados en aquello de la productividad. Sin embargo, tengo un hijo Ingeniero Químico con el que estoy estudiando el asunto. La relación anión-catión, no solo tiene que ver con el estómago de los rumiantes, ni es sólo en los rumiantes. De todas maneras, la marginalidad de mi pregunta se refería a buscar una respuesta que podía ser, por qué no, química de los estudiosos de la materia. Gracias, de todas maneras.
Recomendar
Responder

19 de diciembre de 2006
Quiero felicitar al Dr. Martinez por su excelente articulo, sobre todo porque esta muy completo y porque es un tema que todos los ganaderos deben manejar en sus programas de alimentación.
Con respecto al comentario del Sr. Carlos, tambien estoy de acuerdo, el bienestar animal es un tema que esta en el tapete en estos momentos y no por pura moda sino porque realmente se ha comprobado que al manejar a nuestros animales etologicamente obtenemos mayores beneficios, ganancias y satisfacciones personales. Sobre este tema la Dra Grandin Temple ha publicado mucha información comprobada.
A todos, felicidades y mil bendiciones para este nuevo año.
Recomendar
Responder
19 de diciembre de 2006
Los felicito por el artículo, muy completo. Con respecto al tema que comentaron sobre la felicidad de los animales, lo que nosotros llamamos confort, por ejemplo buena aguada, sombra, buen control sanitario, etc. da muy buenos resultados de ganancia en kg. Es la respuesta que da el animal a las comodidades.
Recomendar
Responder
18 de diciembre de 2006
Personas hay que creen que eso de la felicidad de los animales es poca cosa, que se puede resolver en un solo renglón y se da paso a cosas de más interés como el de producir dinero. Son personas prácticas a las que todo les huele o les hiede a filosófico. Sin embargo es cada vez más evidente que una de las obligaciones del hombre será encargarse de la felicidad de los animales. Acabo de leer un artículo referido a la vida de Teilhard de Chardin, pastor de almas, que se refiere a ello, a través de su concepto sobre la creciente hominización del mundo, es decir, el mundo a imagen y semejanza del hombre. Es cada vez más evidente que las personas adultas besan a los animales y los acarician como si éstos pudieran corresponderles o entender la caricia, lo cual nada tendría de raro, aunque hayan sido anatematizados los animales como irracionales. Incluso se conoce que los horarios de comida de algunos animales domésticos se adecúan a los horarios humanos. Chardin, que además de jesuita fue antropológo, estudió la vida salvaje y las diferencias abismales con la que se pretende cobijarlos. Toros y vacas mientras tanto son privados de sus cuernos, sin que se sepa todavía si, de alguna forma, esto afectará a largo plazo los comportamientos genéticos de los mismos. Se nota si que un animal sin cuernos es más dócil, es casi amaricado aunque, aparentemente, sus cualidades de engendrar y su virilidad no se vean afectadas. Las vacas, ni las gallinas, por ejemplo,pueden ser cortejadas por sus respectivas consortes. ¡Y hay que ver lo que lucha un toro, y los aspavientos de un gallo, para lograr los requiebros de su respectiva amada! Ese ritual erótico, que produce gran cantidad de hormonas, y feromonas, sin las cuales la vida es horrible, ha sido reemplazada por las llamadas pajillas o alguna otra cosa. Esto, que se está convirtiendo en un discurso que algunos mirarían como antitecnológico, no lo es tal. En el centro de la tecnología deberá estar el cultivo del amor y de la felicidad. Esto es cada vez más claro y los que primero lo miren obtendrán las mejores ganancias, y así volver a lo práctico. Así como se está imponiendo lo orgánico, también se impondrán las prácticas de manejo que asuman la Felicidad como una tecnología que trae más beneficios a la humanidad.
Recomendar
Responder
16 de diciembre de 2006
Este artículo más que leerlo hay que rumiarlo, pasarlo por cuatro estómagos. Sin embargo, aunque es muy técnico, luego de tres o cuatro lecturas de los novatos, algo se asimila, aunque sean hierbas. Ignoro si será apto para Navidad. Una pregunta me asalta. Durante todos esos juegos dietéticos ¿la vaca es feliz?. Es decir,tal manipulación o intromisión en sus pautas naturales de alimentación ¿le hace vivir su corta vida más feliz? Se piensa solo en el dinero. Lo digo porque, normalmente, no es preocupación de nadie la felicidad de los animales y, según creo, eso incide mayormente en la calidad de sus productos, uno de ellos, obviamente, el ternero. Lo cual da más dinero. Felicitaciones por la presentación excelente del trabajo. ¡Están contratados! ¡Feliz Navidad! Gracias por tan preciado esfuerzo. ¡Todas las vacas del mundo os agradecerán!
Recomendar
Responder
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar
