La androesterilidad en el mejoramiento genético del sorgo (Sorghum bicolor, L. Moench)
Publicado: 21 de junio de 2022
Por: René Clará Valencia (Coordinador Regional INSTSORMIL y Fitomejorador de Sorgo CENTA) y Nora E. D’Croz-Mason (Adjunct Assistant Professor, University of Nebraska, Lincoln, U.S.A.)
INTRODUCCIÓN
Los métodos convencionales de fitomejoramiento han ofrecido a los Fitomejoradores grandes progresos en el desarrollo de las variedades mejoradas. Sin embargo, sólo una limitada proporción de la variabilidad genética disponible en el sorgo, ha sido explotada. Los adelantos en el mejoramiento de maíz basados en la teoría de la genética cuantitativa han ayudado a entender el uso de las metodologías avanzadas en el mejoramiento de población, utilizando en mayor grado la variabilidad genética. El desarrollo y mejoramiento de poblaciones mediante el uso de la selección recurrente que consiste en ciclos repetidos de selección y recombinación ofrecen la oportunidad de explotar una máxima variabilidad genética, rompiendo así los grupos de ligamientos y liberando la variabilidad oculta para obtener continuas y mejores ganancias genética a un largo plazo (Allard, 1967).
Con la utilización de los genes de esterilidad masculina o androestériles en el sorgo y en otras plantas autógamas, se han podido aplicar las técnicas de poblaciones en forma similar que en el cultivo de maíz. Con las experiencias obtenidas en el Centro Nacional de Tecnología Agropecuaria y Forestal (CENTA) utilizando el gene de esterilidad masculina en sorgo, se han podido romper los grupos de ligamientos como la altura de la planta y la madurez fisiológica, obteniéndose así una diversidad de genotipos con características fenotípicas favorables. También se han obtenido buenos resultados al utilizar genes con esterilidad masculina citoplasmática y genes restauradores de la fertilidad en la producción de híbridos graníferos y forrajeros.
ANDROESTERILIDAD
Al clasificar las plantas por su inhabilidad de producir semillas, es importante hacer distinción entre la esterilidad y la incompatibilidad. Cuando existe un fallo funcional de las anteras o el polen, se denomina esterilidad masculina o androesterilidad. En las plantas androestériles, las flores no producen anteras o polen viable,pero los estigmas funcionan normalmente. Aunque estas flores no pueden ser autopolinizadas, se pueden cruzar con otras fuentes de polen. Esto hace que la androesterilidad sea de utilidad para el fitomejorador. A diferencia, la incompatibilidad es una forma de infertilidad causada por la inhabilidad de las plantas con gametos funcionales, ya sean masculinos o femeninos, de producir semilla cuando sean autopolinizadas o cruzadas. Es un proceso bioquímico bajo un control genético simple. Se presenta en ambos gametos y puede ocurrir en cualquier momento entre la polinización y la fertilización.
Es la esterilidad de los gametos masculinos, se vuelven no funcionales por el efecto de los genes mutantes de los multiples loci que controlan las diferentes etapas vitales para la formación del polen en el núcleo, por los factores citoplasmáticos, o por el efecto combinado de ambos. La androesterilidad no es un mecanismo común para controlar la hibridación en poblaciones naturales; puesto que las plantas androestériles aparecen esporádicamente en poblaciones tanto de especies autógamas como de alógamas (Clará R, 1980).

Foto 1: Flor de sorgo androestéril. Puede notarse que las anteras están atrofiadas y los estigmas son receptivos.
El uso de la androesterilidad, permite a los fitomejoradores eliminar el tedioso proceso de la emasculación en muchas especies autógamas, facilitando de esta manera la producción de híbridos a nivel comercial, lo cual es difícil en las plantas autógamas.
La androesterilidad, según la forma como esté controlada, puede ser: Genética, Citoplasmática y Genética-citoplasmática. Existe otra clase de androesterilidad causada por el medio ambiente, por temperaturas muy bajas, u otros factores abióticos (androesterilidad ecológica), la cual no se tratará en esta publicación.
ANDROESTERILIDAD GENETICA
La androesterilidad genética se manifiesta mediante genes en el núcleo (genes nucleares) que inhiben el desarrollo normal de las anteras y el polen. Esta androesterilidad se ha encontrado en varias especies y está controlada por el gen recesivo “ms”, mientras que el gene dominante “Ms”, produce plantas con anteras y polen normales. Las plantas con en genotipo “ms ms”, son androestériles, mientras que las “Ms ms” o “Ms Ms”son andrpfértiles. Como las plantas androestériles no se pueden mantener por sí mismas deben ser polinizadas por plantas fértiles que lleven, por lo menos, un gen dominante (Ms ms). Si la polinización se lleva a cabo utilizando un gene dominante, (Figura 1), la desendencia de la población será la mitad fértil y la mitad estéril (50% Ms ms:50% ms ms).
En algunas especies se utiliza la androesterilidad genética para la producción comercial de híbridos, sembrando como progenitor femenino una línea heterocigótica (Ms ms) segregando para androesterilidad o una homocigótica (ms ms) y como progenitor masculino una línea homocigótica dominante (Ms Ms) o heterocigótica (Ms ms); al momento de inicio de la floración y antes de iniciarse la antesis, se identifican y se eliminan las plantas fértiles en el progenitor femenino.
Algunas especies poseen características fenotípicas asociadas con la androesterilidad genética, las cuales sirven para identificar las plantas androestériles (Judía de Lima).
El problema en la producción de híbridos comerciales no es solamente el costo elevado de la semilla sino también la dificultad en identificar las plantas androfértiles en los surcos del progenitor femeniono. En los cultivos de tomate, judías y cebada la producción de semilla en las plantas androestériles es escasa, debido a la falta de buenos polinizadores. En el tomate, posiblemente se puede obtener un mejor rendimiento de semilla bajo condiciones ambientales que favorezcan la polinización.

En la mayoría de las especies autógamas donde se presenta esta clase de androesterilidad, su principal uso es en los programas de mejoramiento es para aumentar la variabilidad mediante la recombinación al azar con diferentes progenitores masculinos.
ANDROESTERILIDAD CITOPLASMATICA
Esta clase de esterilidad masculina está controlada por factores citoplasmáticos. Las plantas androestériles (ms ms) poseen citoplasma estéril (S) y genes homocigóticos recesivos para la fertilidad en el núcleo (ms ms)S. Estas plantas producen semilla y mantienen su esterilidad masculina cuando se polinizan con plantas de citoplasma fértil (F) y núcleo con genes recesivos para la fertilidad (ms ms)F. Las primeras se conocen como líneas “A”, las segundas como línes “B” o mantenedoras de la androesterilidad.
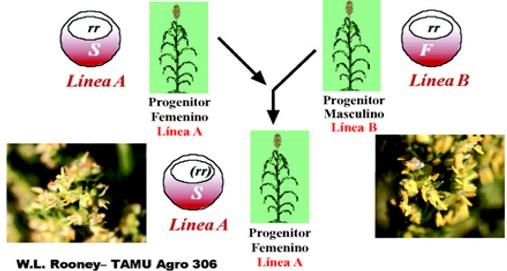
Fig. 2. Mantenimiento de semilla de la línea “A”
En la figura 2 se puede observar que la diferencia de los progenitores es solamente en el citoplasma y la descendencia lleva siempre el citoplasma (ms ms)S, o sea con dominancia del citoplasma estéril (S), lo que también se conoce como herencia materna o matroclinia. Los híbridos obtenidos mediante esta metodología son androestériles y no producen semilla; por lo tanto no es importante en cultivos donde el producto comercial es la semilla.
La androesterilidad citoplasmática es bastante utilizada en plantas ornamentales, debido a que los híbridos androestériles producen flores más hermosas y se mantienen frescas mucho más tiempo que las plantas androfértiles dentro de la misma especie.
El gene de esterilidad masculina, se conoce como “ms”, el androestéril citoplasmático como “msc”. Sin embargo para mejor explicación, también se utiliza RR y rr para denominar lo genes dominantes y recesivos, de la androesterilidad (Allard, 1967).
ANDROESTERILIDAD GENETICA-CITOPLASMATICA
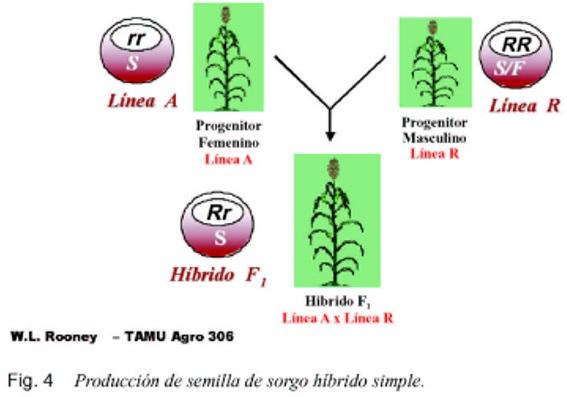
Es una interacción entre el núcleo y el citoplasma que produce plantas androestériles y fértiles. Se restaura la fertilidad en la F1 cuando se cruzan plantas con citoplasma estéril y genes homocióticos recesivos para la fertilidad (rr)S, con plantas de citoplasma estéril o fértil y genes homocigóticos dominantes en el núcleo (RR)_. Estos últimos genes tienen la capacidad de restaurar la fertilidad del polen en el citoplasrna androestéril, y se conocen como “Restauradores” y las plantas como líneas “R”.
Esta clase de esterilidad fué reportada por Stephens y Holland en 1954 en el cultivo del sorgo, al transferir cromosomas de Kafir en el citoplasma de Milo y obteniendo plantas androestériles y fértiles en la progenie de la F2..
Allard R.W. 1967, menciona que este tipo de androesterilidad fue utilizada primeramente en cebolla por Jones y Davis (1944), cuando encontró una planta en la variedad Italian Red que era
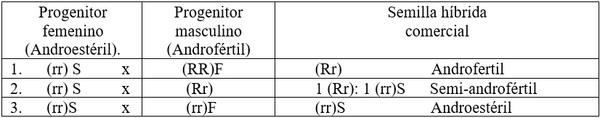
Cuadro 1. Métodos de producción de semilla híbrida comercial.
completamente androestéril. Ellos la cruzaron con varios tipos de plantas androfértiles y se obtuvo tres tipos de descendencia: un primer tipo fue androfértil completamente, un segundo produjo plantas androfértiles y androestértiles en proporción 1:1 y un tercero fue completamente androestéril (Cuadro 1). Algunos tipos de citoplasmas interaccionan de forma diferente con el núcleo, por esta razón algunas líneas “R” se vuelven líneas “B” cruzándolas con otros citoplasmas.
Estos experimentos dieron la pauta para la producción comercial de semilla híbrida en varias especies autógamas, como cebolla, ajonjolí, sorgo, arroz, etc.- Este tipo de androesterilidad también se usa en maíz, para disminuir el alto costo del desespigue. El éxito obtenido utilizando este tipo de androesterilidad se debe al bajo costo de la semilla híbrida y a su facilidad en el proceso de producción en el campo..
Si el producto comercial es la semilla, el método adecuado es el primero, pero si el producto comercial es la parte vegetativa pueden utilizarse los tres métodos.
La androesterilidad genética-citoplasmática se utiliza para producir híbridos simples (figura 3), triples (figura 4) y dobles (figura 5).

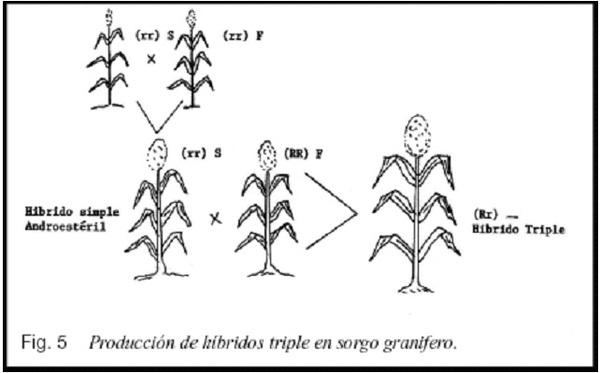
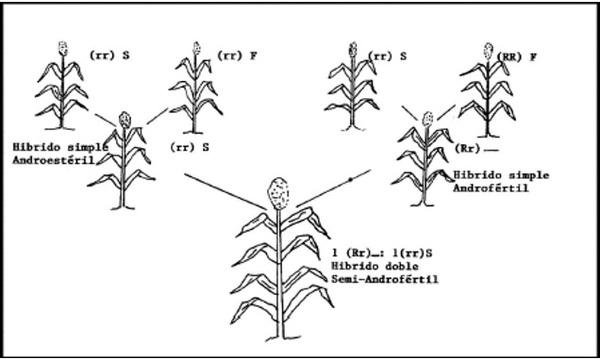
La Fig. 5, enseña la forma como de producir semilla en un híbrido doble, pero nótese que el híbrido produce plantas androestériles y androfértiles en proporción 1:1, lo cual podría reducir el rendimiento por una deficiente polinización, pero si el producto comercial fuera la parte vegetativa, esto no sería problema.
Los híbridos dobles son de uso más frecuente en maíz; en sorgo no se utilizan.
ANDROESTERILIZACION DE LINEAS “B”
Cuando se tiene un grupo de líneas nuevas en las que se desconoce la fertilización del polen, se cruzan todas con una línea androestéril (A) y luego se siembran todas las F1. Se obtienen tres tipos de progenies: androfértil, semifértil y androestéril (Cuadro 1). En sorgo para grano solo interesa la primera y la tercera. En las F1 androestériles las líneas utilizadas como progenitor masculino le llaman lineas mantenedoras o líneas “B” o sea (rr)F con citoplasma fértil y las F1 androfértiles son líneas “R” o restauradoras (RR)_, con genes dominantes para la fertilidad en el núcleo.
Las mejores líneas “B” es importante androesterilizarlas, o sea transferir los cromosomas de la línea “B” al citoplasma de la línea “A” (rr)S. Esto se realiza mediante selección apareada de progenie en un proceso de retrocruzamiento (House, 1985).

Foto No. 3. Androesterilización de lineas “B”. Estac. Experimental de Santa Cruz Porrillo (CENTA), 1977
El proceso de androesterilización puede comenzar antes o después que la línea sea homocigótica y mientras se desarrolla, prosigue la selección en busca de caracteres que converjan en la línea “B”, hasta obtener mas de un 98% de todos los caracteres de la línea “B”, pero con la diferencia que no produce polen viable (Fig. 6).
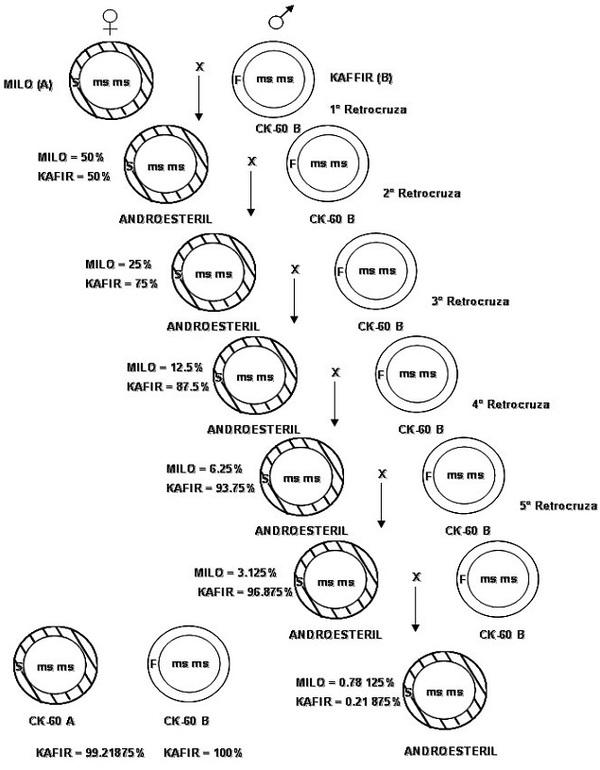
Figura 6. Proceso de esterilización en sorgo. utilizando la FF androestéril del cruzamiento Milo X Kafir, bajo seis generaciones de retrocruzamiento con Kafir. Depués del sexto retrocruzamiento casi todos los genes del núcleo (99.21875%) son reconstituidos en la progenie, conservándose el gene msms intacto (Nagur, 1981).
DESARROLLO DE POBLACIONES RANDOMIZADAS
A diferencia de los métodos convencionales de fitomejoramiento, el mejoramiento de poblaciones por medio de selección recurrente es a largo plazo, donde la ganancia genética es baja pero el sistema mantiene una variabilidad genética más amplia y durante un período más largo; además, los frecuentes intercruzamientos darán origen a nuevas recombinaciones que aumentan las oportunidades de selección a través de cada ciclo. Siguiendo al descubrimiento de la esterilidad masculina (ms) y al desarrollo de la teoría genética cuantitativa en maíz, nuevas poblaciones randomizadas, han sido desarrolladas en muchos programas de fitomejoramiento.
El desarrollo de las poblaciones randomizadas involucra tres pasos básicos:
a) Identificación o selección de progenitores adecuados.
b) Incorporación del gene de esterilidad masculina (ms).
c) Intercruzamientos o recombinación de los progenitores.
La selección de los progenitores para formar la población básica es muy crítica y tiene sus implicaciones en el valor de la población, en la selección y en el avance del mejoramiento. El objetivo de la selección recurrente es mejorar el comportamiento de la población para una o mas características como: rendimiento y sus componentes, factores de estabilidad, resistencia a plagas y enfermedades y características de calidad de grano. Los progenitores deben de estar representados con líneas de diferente origen y buena aptitud combinatoria para maximizar la diversidad genética.
El número de progenitores varía según los objetivos; una población con menos de 10 progenitores no proporciona el alcance suficiente para una selección simultánea. Demasiados progenitores resultan en una población muy diversa, pero con bajo potencial de rendimiento en la población (Cuadro 2).
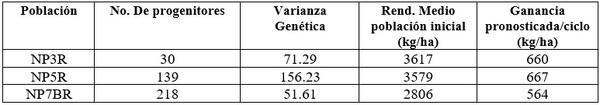
Cuadro 2. Número de progenitores, variabilidad genética, rendimiento promedio y las ganancias pronosticadas por ciclo de selección para tres poblaciones
Es importante incorporar suficiente variabilidad en la población, porque es la base para el éxito de cualquier programa de fitomejoramiento. No existen reportes sobre el número exacto de progenitores que deban usarse.
En el Cuadro 2 se ve que aunque el número máximo de progenitores estuvo involucrado en la población NP7BR la varianza genética fue más elevada en la NP5R. El rendimiento medio de grano antes de la selección fue el más bajo para NP7BR y más elevado en NP3R.
La introducción de genes con esterilidad masculina se realiza por retrocruzamiento. Por lo menos en una etapa durante la retrocruza, las cruzas deben hacerse usando los progenitores recurrentes como hembra, a fin de tener variabilidad de factores citoplasmáticos en la población. Generalmente de 2 a 3 retrocruzas seguidos por selección, son suficientes para alcanzar los objetivos, particularmente cuando la fuente del gen androestéril tiene antecedentes élites.
Cuando se agregan nuevas entradas o progenitores a la población se debe tener cuidado que el compuesto permanezca balanceado. No debe mezclarse por igual el número de semillas, tanto del compuesto como de la entrada nueva que se desea introducir. Si el compuesto está en sus primeras etapas de desarrollo, posiblemente de 5 a 10 gramos de semilla de la entrada nueva, pueden mezclarse con 1000 gramos del compuesto. Si este ya ha sido avanzado por varios ciclos de selección, entonces será conveniente cruzar y retrocruzar la nueva entrada por el compuesto.
También es posible desarrollar un “Side Car” donde el compuesto original es cruzado sobre la nueva fuente y retrocruzado como el progenitor recurrente. La selección es continuada en el mejoramiento original y en el “Side Car”, de manera que ambas poblaciones son mejoradas simultáneamente. Lo mejor del compuesto original es conservado con esta técnica.
La recombinación de los progenitores dentro de la población se realiza en lotes aislados, dejando que las plantas androestériles que segregan en la S1, se polinicen con las plantas fértiles heterocigóticas y homocigóticas dominantes. Son suficientes de 2 a 3 ciclos de recombinación, para que la población sea expuesta a cualquier proceso de selección.

Foto 4. Población ESPF-1 en proceso de formación, CENTA, El Salvador, 1978.
METODOS DE MEJORAMIENTO INTRAPOBLACIONAL
Una vez la población ha sido formada, pueden usarse los siguientes métodos de selección:
Selección masal
Es un método fácil y sencillo para poblaciones heterogéneas y de caracteres con alto coeficiente de heredabilidad. Cada ciclo representa una generación, mientras que otros sistemas requieren más de uno. Un gran lotede germoplasma puede ser muestreado; en otros sistemas donde se requieren pruebas de rendimiento, el número de muestras para ser evaluadas es más limitado.
Este método consiste en: a) la selección de plantas individuales con base en su fenotipo y b) la mezcla de cantidades iguales de semilla de cada planta seleccionada para formar el próximo ciclo de selección. En sorgo, donde la selección es basada en plantas androestériles, se identifican las hembras durante la floración y se seleccionan durante la madurez. Estas plantas son polinizadas por el polen proveniente de plantas con buenas o malas características involucradas en la población (House L.R. 1985). La desventaja de la población masal es que no controla el origen del polen ni su contribución en las progenies, por esta razón la heredabilidad se reduce a la mitad. Si la selección se efectúa en plantas fértiles, se necesitan dos generaciones por ciclo, un ciclo adicional para la recombinación. Doggett, H. and Eberhart, S.A. 1968, propusieron alternar la selección de las plantas fértiles y androestériles. En este sistema, la selección de las plantas androestériles se hacen en la primera siembra y las de las fértiles en la siguiente y así sucesivamente. Una generación es esencialmente de selección y la otra de recombinación.
En el CENTA al aplicar la selección masal en la población ESPF-1 (ms3) con androesterilidad genética, se obtuvo buen progreso durante tres primeras generaciones, pero posteriormente la población declinó en genotipos con poco vigor y con una media de rendimiento inferior a la inicial.
Selección de familias de medios hermanos
Este es otro sistema fácil de usar en sorgo, donde la androesterilidad genética ha sido incorporada y toma dos generaciones por ciclo. En un lote aislado las plantas androestériles se marcan durante la floración y se dejan polinizar libremente. Durante la cosecha, cada panoja seleccionada se desgrana individualmente. La semilla de cada panoja representa una entrada o familia que debe ser evaluada en un ensayo de rendimiento. El resto de la familia se guarda para ser utilizada una vez se obtengan los ensayos de rendimiento. Estas familias se siembran en ensayos replicados durante la estación principal. Generalmente se utilizan más de 250 entradas (cada entrada es una panoja de planta androestéril).
Las mejores entradas de los ensayos de rendimiento se seleccionan y se mezcla la semilla que se guardó y se siembra para comenzar un nuevo ciclo de recombinación. Nuevamente, las plantas androestériles son identificadas durante la floración y cosechadas individualmente para formar el siguiente ciclo de selección.
El método se llama “Selección de familia de medios hermanos” porque la selección se realiza en un solo progenitor, en este caso el femenino androesteril, que es polinizado al azar por progenitores no conocidos.
Selección de familias de hermanos completos
El procedimiento es igual al de medios hermanos, excepto que la recombinación se realiza en cruzamientos apareados entre plantas fértiles y androestériles, o sea que hay control en ambos progenitores. Este sistema de selección es llamado de hermanos completos y son también evaluados en ensayos de rendimiento. Es difícil de usarlo en la práctica, cuando se obtiene insuficiente semilla en los cruzamientos para montar varios ensayos.
Selección Familias S1
Se refiere a la progenie de una población libre que ha sido polinizada. Pueden utilizarse progenies segregantes para androesterilidad o progenies con fertilidad uniforme.
Este sistema requiere tres generaciones y es el más efectivo cuando se pueden realizar las tres cosechas en un año. Un gran número de plantas androestériles de polinización abierta son cosechadas y sembradas en panoja por surco en un vivero sin replicar. Cerca del 50% de las líneas pueden ser eliminadas con base en sus características agronómicas. La mejor planta fértil se selecciona de cada familia de medios hermanos seleccionada. Estas plantas proveerán semilla para la evaluación de la progenie S1.
Un mínimo de 200 a 250 líneas S1 son evaluadas en ensayos replicados preferiblemente en varias localidades. Las mejores (10-20%) entradas son seleccionadas, y el remanente de esa semilla es usado para recombinar la población y el ciclo se repite.
Juida Jan Orn et al 1976 (mencionado por Lukhele, P. and Obilana A.T. 1980) al comparar los métodos de selección masal, medios hermanos y Familias S1 para mejorar la población de sorgo NP3R, encontraron que la respuesta predicha para el rendimiento de grano fue máxima en las familias de las S1 evaluadas. Doggett (1972) reportó un incremento promedio de 25% en rendimiento de grano en cuatro poblaciones después de un ciclo de selección. El máximo rendimiento (33%) fue observado en la población PRS1.
Familias S2
Este sistema (Figura 8) es una extensión adicional del S1. Las líneas S2 (plantas fértiles provenientes de las líneas S1) son evaluadas en ensayos replicados en vez de líneas S1. Se necesitan cuatro generaciones para completar un ciclo. Este sistema es muy efectivo en la eliminación de genes indeseables de las poblaciones e incrementa la varianza genética aditiva.
Debido a que se realizan dos autofecundaciones sucesivas, la frecuencia de las plantas androestériles se reducen bastante, por lo tanto se hace necesario realizar polinizaciones de plantas hermanas (Sibbing) de cada progenie S2en viveros separados. La semilla de los Sibbing de las líneas seleccionadas se utiliza para la recombinación de la población (Clará R. 1980).

Figura 8. Método de mejoramiento de familia S2
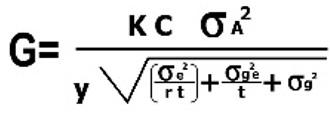
Una vez que una población se ha desarrollado bajo los distintos métodos de selección previamente mencionados, la ganancia genética puede obtenerse mediante la fórmula siguiente:
Donde:
K= Indice de selección, diferencial estandarizado.
C= Control parental, es 1 cuando la unidad de selección es la misma que la unidad de recombinación, ambos padres son seleccionados; es igual a ½ cuando solo el progenitor femenino es seleccionado.
σ2A= Varianza genética aditiva y= Número de ciclos por año.
σ2e= Varianza del error experimental
σ2ge= Varianza de la interacción genotipo-ambiente.
σ2g= Varianza genética r= Número de repeticiones t= número de localidades
C= Control parental, es 1 cuando la unidad de selección es la misma que la unidad de recombinación, ambos padres son seleccionados; es igual a ½ cuando solo el progenitor femenino es seleccionado.
σ2A= Varianza genética aditiva y= Número de ciclos por año.
σ2e= Varianza del error experimental
σ2ge= Varianza de la interacción genotipo-ambiente.
σ2g= Varianza genética r= Número de repeticiones t= número de localidades
Esta fórmula revela que la ganancia genética debido a la selección puede ser aumentada de varias formas.
1. Intensidad de selección
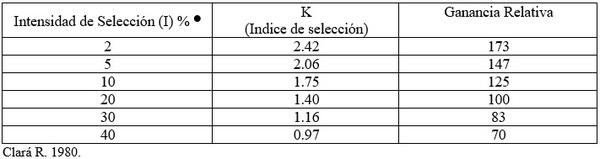
La ganancia puede ser aumentada teniendo una intensidad más elevada de selección (Cuadro 3).
La ganancia puede ser duplicada si se usa con una intensidad de selección de 5% en vez de 40%. Sin embargo, a medida que se aumenta la intensidad de selección, el número de familias evaluadas debe ser aumentado, de manera que el tamaño de la población efectiva sea mantenido lo suficientemente grande, para evitar la pérdida de los alelos favorables, a través de fijación al azar (erosión genética).
Cuadro 3. Relación entre la intensidad de selección (i) y la ganancia genética.
2. Efectos de la selección en la varianza fenotípica
El aumento de la heredabilidad (h2 =σ2A/ σ2P) por reducción de la varianza fenotípica es otro medio de aumentar la ganancia de la selección. La selección reduce la varianza fenotípica, genotípica y la heredabilidad en las progenies, durante el primer ciclo. Como la varianza fenotípica incluye la varianza del error (σ2e), la interacción genotipo-ambiente (σ2ge) y la genética (σ2g), las técnicas mejoradas en parcelas de ensayos reducirán la varianza del error y eficientes diseños estadísticos experimentales que incluyan suficiente número de ambientes y repeticiones, reducirán la varianza fenotípica.
Exkebil (mencionado por House L.R. 1985), encontró mayor ganancia en el rendimiento del sorgo utilizando dos repeticiones durante dos años.
3. Métodos de mejoramiento
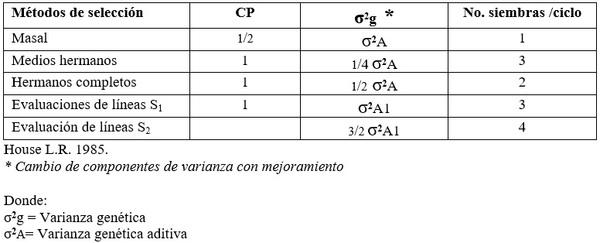
También puede ser aumentada la varianza genética aditiva entre las familias, al cambiar el control parental o reduciendo el número de años por ciclo (Cuadro 4).
Cuadro 4. Control parental, varianza genética aditiva y número de siembras por ciclo para diferentes métodos de selección
METODO DE MEJORAMIENTO INTERPOBLACIONAL
Los métodos de fitomejoramiento interpoblacional son usados cuando el objetivo principal es el desarrollo de híbridos. El sistema se utiliza en cultivos donde la dominancia, epistasis y la acción genética aditiva son importantes. En sorgo la acción aditiva del gene es más predominante que la dominancia o la sobre dominancia. Sin embargo, hay reportes donde la dominancia ha sido encontrada considerablemente alta. Desafortunadamente, los componentes apropiados de la varianza no están disponibles para comparar las ganancias esperadas, provenientes de los métodos de mejoramiento interpoblacional. La selección recurrente recíproca, donde se mejoran dos poblaciones simultáneamente por su aptitud combinatoria general y específica, utilizando cada población como probador, es tediosa para seguir en sorgo. Russell y Eberhart, 1975 (mencionado por Obilana 1980) sugirieron una modificación a este sistema de selección e indicaron que se obtiene una variación mas grande entre las familias cuando una línea pura de la población recíproca se utiliza como probador, esperándose que produzca un mayor progreso en la selección que cuando la población recíproca se usa como probador.
Aunque el uso de un probador de línea pura fue propuesto inicialmente para mejorar la población de genes con efectos dominantes, los resultados de Horner et al (mencionado por Lukhele P. y Obilana A.T. 1980), indican que los probadores de línea pura son efectivos en seleccionar genes con efectos aditivos, porque muchos de sus loci son homocigóticos para los alelos recesivos. Ellos también sugirieron que las poblaciones por selección recurrente recíproca, usando un probador estrecho pueden combinar bien con líneas no relacionadas, diferentes de las usadas como probador.
DISEÑOS ESTADÍSTICOS EXPERIMENTALES PARA EVALUACIÓN DE LAS POBLACIONES
En un programa de selección recurrrente un gran número de familias provenientes de varias poblaciones, generan muchas parcelas las cuales deben ser evaluadas . Sin embargo, es indispensable este gran número de familias para poder estimar adecuadamente las medias, las varianzas, las heredabilidades y predecir la ganancia genética en estudios de genética cuantitativa. La evaluación de estos experimentos es compleja, requiere atención especial, e involucra diseños experimentales que no son muy comunes.
La demanda por la adquisición de la tierra, el tiempo, y la mano de obra, entre otras cosas es díficil cuando se conducen muchos experimentos. Estas demandas se reducen significativamente al seguir la técnica de Ross y Gardner (1983), quienes para evaluar las poblaciones de sorgo sugirieron utilizar parcelas de un surco de 7,6 m en vez de dos o más surcos, cosechando solamente 4,4 m en dicho surco. Incluyendo las calles, en una hectárea se pueden acomodar 1430 parcelas de un surco cada una.
El número apropiado de familias para evaluar fluctúa entre los 100 y los 200. Menos de 100 individuos no estiman bien los parámetros genéticos y más de 200, aunque dan excelentes resultados son difíciles de manejar. La desventaja en evaluar menos de 200 familias depende en la intensidad de la selección que se aplique, la cual seguramente no proveerá un número adecuado de familias para ser recombinadas, promoviendo de esta manera un alto nivel de consanguinidad. Se puede ganar precisión en la evaluación de las familias al aumentar el número de repeticiones, pero al aumentar las repeticiones se incrementa el costo de los experimentos. En general, se recomienda aumentar los sitios de pruebas o localidades (Ross y Gardner, 1983)
Como los fitomejoradores están interesados en la interacción genotipo x ambiente, las familias deben ser evaluadas en más de una localidad y por más de un año. Cuando se evaluan diferentes años y diferentes localidades, el procedimiento correcto con cualquier diseño experimental es calcular ambas fuentes de variación, la interacción de las familias con ambos, años y localidades, y la interacción año x localidad (Ross y Gardner, 1983). Si las localidades no son las mismas cada año, entonces se consideran las localidades como efectos anidados dentro de los años. En este caso, no hay interacción año x localidad y tan solo se estiman las interacciones de familias x años y localidades x años. Para este caso, se recomienda tratar todos los experimentos localidad - año como ambientes separados. Esto producirá solamente interacciones familia x ambiente, simplificando así la estimación de los componentes de varianza genética y la de genotipo x ambiente. De esta manera, se asume que cada experimento localidad - año representa una muestra al azar del ambiente en el cual se realiza el mejoramiento (Ross y Gardner, 1983)
Los diseños estadísticos experimentales para evaluar familias provenientes de un programa de selección recurrente aunque llevan muchas entradas y pocas repeticiones, estiman adecuadamente los parámetros que se miden. Algunos diseños dan mejores resultados que otros.
Los estadistas evalúan la eficiencia de un método basado en el promedio de la varianza del error de las entradas, mientras que los genetistas, están mas interesados en la ganancia debido a la selección o sea la disminución del cuadrado medio del error.
El diseño experimental de bloques completamente al azar podria utilizarse para evaluar poblaciones, pero una vez las entradas pasan de 25, el error experimental tiende a aumentar, lo cual no es satisfactorio.
El diseño experimental de lattice, es excelente cuando existe un gran número de entradas. Jan-orn et al. (1976) con un lattice de 14x14 obtuvieron una mejor ganancia genética en 196 familias S1 de sorgo, que al utilizar bloques completamente al azar. La desventaja de los lattices radica en el número de tratamientos que debe ser siempre un cuadrado exacto (n2) o un n (n-1) de entradas para lattices rectangulares (Cochran y Cox , 1992). Cuando hay errores en la siembra o más adelante si hay perdidas de parcelas, se pierde la eficiencia del lattice, teniendo que analizarse como un bloque completamente al azar.
Para superar el uso de bloques muy grandes, Yates en 1936 creó un diseño pseudo-factorial o de bloques incompletos. En este arreglo, las variedades se dividen en juegos o grupos para ser comparados en más de una forma. Por ejemplo, 100 variedades denominadas del 1 al 100 pueden ser divididas en juegos o grupos de 10, en dos formas: el primer grupo de 10 por las variedades del 1 al 10, 11-20, 21-30 ... 91-100, y el segundo grupo de 10, por las variedades 1, 11, 21, 31,..91; 2, 12, 22,...92 etc. Cada grupo de 10 se puede acomodar en el campo en uno o más bloques randomizados o en cuadrados latinos de acuerdo al número de repeticiones que sean posibles. Lo que imposibilita el uso de los cuadrados latinos en vez de los bloques al azar es el gran número de repeticiones requeridas.
La información sobre la diferencia entre las variedades 1 y 2 puede obtenerse de dos maneras. La primera, proveniente de la comparación dentro del grupo 1-10 y la segunda utilizando el promedio del grupo 11, 21...91 para la variedad 1 y el promedio de 12, 22, 32...92 para la variedad 2, la diferencia entre las dos medias siendo determinada por la comparación dentro del primer juego de grupos (Yates, 1936)
Schutz y Cockerham (1966) compararon 3 diseños de bloques incompletos en 256 tratamientos con 16 bloques y 2 repeticiones y también evaluaron la ventaja de usar testigos o controles dentro de estos diseños los cuales designaron: B - bloques en (dentro de) repeticiones, R - repeticiones en (dentro de) bloques, y S - lattices. Los autores concluyeron que el diseño B era casi igual o superior al R y que no era ventajoso incluir testigos a costa de las variedades a probar.
El número total de tratamientos o entradas para todos diseños de bloques incompletos es T = pb donde p representa el número de entradas por bloque, y b el número de bloques por repetición. Conr repeticiones, el total de unidades experimentales es entonces T= = rT. En el diseño de lattices, T = p2 y r = 2. En los diseños B y R las mismas entradas p deben aparecer siempre juntas en el bloque incompleto y en cada repetición. La diferencia entre estos dos diseños es la manera en la cual un bloque incompleto (grupo) de entradas es repetido en el campo.
Para el diseño B,r repeticiones completas de T parcelas contiguas son subdivididas en b bloques de p parcelas cada una, y los grupos de tratamientos asignados al azar a los bloques, en cada repetición. En el diseño R, el campo experimental es inicialmente subdividido en grupos rpde parcelas contiguas, dentro de los cuales un bloque incompleto de p entradas es repetido r veces. El diseño S, es similar al B, con r=2 , pero en los lattices cada entrada aparece con un grupo diferente de entradas en cada uno de los dos bloques incompletos donde ocurren. (Schutz y Cokerham, 1966).
Los modelos para los tres diseños son:
Yi j k = μ + r i + g i j + P j k + e i j k ( B )
Donde μ es la media, jk simboliza la entrada y jel bloque en la enesima repetición (i= 1,2 ...,r; j= 1,2,...,b; y k= 1, 2,...., p ). Los efectos ambientales vienen a ser r para repeticiones, g para bloques en repetición, e para el error y P el efecto de las entradas. Los efectos, se asume que son al azar, no correlacionados y con varianzas para cada categoria de: σ2r, σ2g,σ2 y σ2 p respectivamente.
Yi j k = μ + G j + R j i + P j k + e i j k ( R )
Para este diseño,idenota la repetición en el grupo j. Los effectos ambientales cambian, siendo G el efecto para el grupo de rrepeticiones contiguas de un grupo de entradas en un bloque incompleto y R los bloques incompletos repetidos dentro de los grupos. Sus varianzas son σ2G y σ2R,
El modelo para lattices ( S ) es muy similar al de bloques en (dentro de) repeticiones ( B ):
Yi j k = μ + ri + g i j + P īj k + e i j k
P īj kse interpreta como P j k= P j k
Repeticiones en (o dentro de) bloques
El diseño experimental repeticiones en (o dentro de) bloques fué utilizado inicialmente por Comstock y Robinson (1948) cuando evaluaron 192 progenies de maiz, obtenidas al recombinar 48 machos con 192 hembras. Las cuatro progenies de un grupo tenían un padre en común pero una madre diferente. En el campo, cada bloque tenía 32 parcelas. Las progenies se dividieron en 12 juegos o grupos de 4 machos cada uno y cada grupo de 16 progenies fué asignado a un bloque y repetido 2 veces en el (o dentro del) bloque. Las 16 progenies se volvieron a randomizar dentro de cada una de las dos repeticiones. Cuando se desea que los componentes genéticos de las varianzas estimen la heredabilidad, las correlaciones genéticas y la predicción del progreso esperado por la selección, se logra una mayor precisión al agrupar al azar las familias escogida en bloques incompletos, los cuales permanecen iguales de repetición en repetición (Comstock y Robinson 1948).
Ross et al. (1981) utilizaron repeticiones en(dentro de) bloques para evaluar 200 familias S1 en una población de sorgo. El Cuadro A enseña el analisis de varianza en a) una localidad, 2 repeticiones y b) dos localidades, 2 repeticiones para 200 familias divididas al azar en 10 grupos de 20 familias, donde cada grupo permanece unido en ambas repeticiones, y ambas repeticiones ocurren en el mismo bloque. La variabilidad del suelo la cual contribuye al error experimental dentro de una replicación y dentro de un bloque, se reduce notablemente. Cada bloque es esencialmente un pequeño experimento de bloques al azar involucrando un juego de familias randomizado de una población.
En repeticiones en (dentro de) bloques, la media de cada bloque estima el promedio de la población, y la variación entre las familias y la varianza del error en cada bloque son las estimaciones de las mismas varianzas de la población. Por lo tanto, las sumas de los cuadrados, y los grados de libertad para las repeticiones en bloques, las familias en bloques, y el error son aditivas a traves de los bloques (Cuadro A). Al remover la variación entre los bloques y entre las repeticiones dentro de los bloques se provee con una eficiencia más grande que cuando se usa el diseño de bloques completamente al azar para estimar las varianzas (genéticas) de las familias. (Ross y Gardner, 1983).
Ventajas
El diseño de bloques en (dentro de) repeticiones tiene la ventaja de que al perderse un bloque, tan solo se pierden unas pocas entradas en ambas repeticiones en vez de la perdida parcial de una repetición con muchas entradas. Este diseño es utilizado para experimentos en el invernadero, cámaras de crecimiento, etc, porque los potes, o cajas que representan los grupos o bloques pueden descartarse sin perder una repetición entera.
Bloques en (o dentro de ) repeticiones
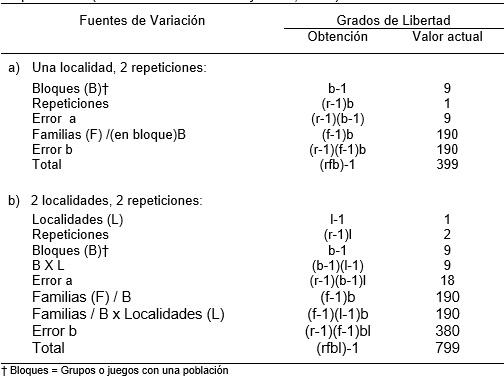
El diseño de bloques en (o dentro) repeticiones está basado en diseños delineados por Yates (1936) para la evaluación de un gran número de entradas en bloques incompletos parcialmente balanceados. Eckebil et al, (1977) y Ross et al. (1983) utilizaron este diseño en 200 familias S1 en tres poblaciones de sorgo donde las entradas por grupo y los grupos por bloque permanecieron juntas en la segunda repetición, donde fueron nuevamente randomizadas. En este estudio, 200 familias fueron divididas en 10 grupos de 20 cada una. Las 20 familias en cada bloque permanecieron juntas en cada repetición teniendo así 10 bloques por repetición. El Cuadro B enseña el analisis de varianza para este diseño en a) una localidad, 2 repeticiones y b) 2 localidades, 2 repeticiones.
Ventajas
La obtención de los componentes de varianza para la familias, y las familias x localidades es la misma para ambos diseños. Sin embargo, los bloques en (dentro de) repeticiones no dan tan buen estimado de estos componentes como el de repeticiones en (dentro de) bloques porque cada familia está más separada en las repeticiones.
Este diseño es muy práctico cuando los tratamientos como fertilizantes, irrigaciones, inoculaciones de enfermedades, e infestaciones de insectos se pueden sobreponer en todas las repeticiones del experimento. Sin embargo, el riesgo es más grande porque se reduce su eficiencia con la pérdida parcial de una repetición, especialmente cuando solo se utilizan dos repeticiones.
EVALUACION SIMULTANEA DE DOS POBLACIONES
La evaluación simultanea de 2 poblaciones en un programa de selección recurrente presenta una situación bastante compleja, por la adición de más fuentes de variación para el analisis estadistico. Además, debe tenerse en cuenta que los efectos ya no son solamente al azar, sino que además existen efectos fijos, y la prueba de F viene a ser muy compleja. Se pueden utilizar ambos diseños de bloques incompletos pero con algunas variaciones en las cuales, para el caso de repeticiones en (dentro de) bloques, los bloques están divididos en repeticiones con un juego de familias proveniente de cada población asignadas al azar en cada repetición. Por ejemplo, un bloque puede tener 2 repeticiones y cada repetición tiene un juego de 20 familias de cada una de las dos poblaciones. La randomización se utiliza todo el tiempo, con la excepción de que las familias de una población en un juego, permanecen juntas en ambas repeticiones, y un bloque tiene el mismo grupo de poblaciones cuando se repite el experimento. Existen muchas variaciones, como dividir cada bloque en poblaciones y subdividir las poblaciones dentro de las repeticiones, cambiandose asi el analísis de varianza (Ross y Gardner, 1983).
En el diseño de bloques en (dentro de) repeticiones un bloque contiene un juego de 20 familias de cada población para un total de 10 bloques por repetición. Se randomizan las familias en los juegos, los juegos en los bloques, y los bloques en las repeticiones, recordando que un juego y un bloque deben tener siempre el mismo germoplasma en ambas repeticiones y en ambos ambientes.
Cuadro A. Analisis de varianza para una población de 200 familias divididas en 10 juegos de 20 familias en a) 1 localidad y 2 repeticiones y b) 2 localidades, 2 repeticiones utilizando el diseño de repeticiones en (dentro de) bloques (Modificado de Gardner y Ross, 1983).

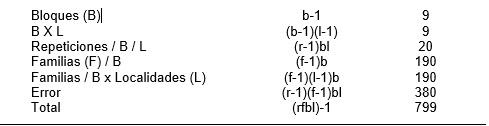
Cuadro B. Analisis de varianza para una población de 200 familias divididas en 10 juegos de 20 familias en a) 1 localidad, 2 repeticiones y b) 2 localidades, 2 repeticiones utilizando el diseño de bloques en (dentro de) repeticiones (Modificado de Gardner y Ross, 1983).
Cuadro C. Esquemas de un diseño aumentado utilizando bloques completamente al azar (Modificado de Federer, 1961; 1975)
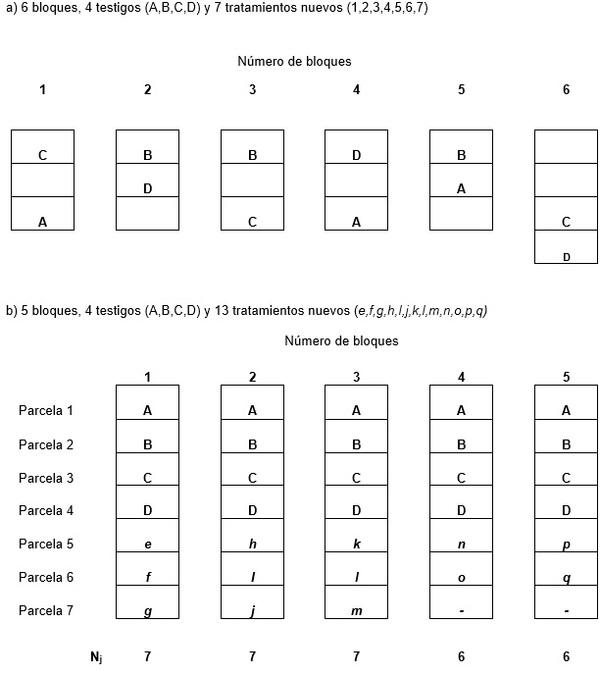
DISEÑOS EXPERIMENTALES SIN REPETICIONES
Existen ocasiones donde no se posee suficiente cantidad del material (semilla) para ser evaluado y es díficil usar dos repeticiones . Muchísimos programas de mejoramiento evaluan las nuevas lineas como hibridos, o poblaciones en desarrollo en parcelas con una sola repetición, y utilizan métodos menos complicados. Federer, 1956, 1961, 1975, sugirió el uso del diseño aumentado (Augmented or Hoonuiaku design)
Un diseño experimental aumentado es un diseño normal con tratamientos adicionales (nuevas lineas, nuevas variedades, o nuevos hibridos , etc) en el bloque completo, el bloque incompleto, el surco, o la columna, etc. Aunque, solamente una parcela de una nueva linea o una población es evaluada en una localidad, estos nuevos tratamientos pueden ser sembrados en diferentes localidades, utilizando de 3 a 5 hibridos, o variedades comerciales o adaptadas como testigos los cuales deben ser repetidos r veces en cada localidad. Es ventajoso incluir estos testigos (3 a 5) en cada bloque y analizar sus datos como un experimento normal (de más de una repetición) para obtener un estimado del error experimental o el cuadrado medio del error. El Cuadro C enseña dos esquemas que se emplean cuando se tienen a) r = 6 bloques, vr= 4 hibridos o variedades comerciales (A, B, C, D) y v1 = 7 nuevos tratamientos o nuevas variedades, poblaciones, o hibridos (1, 2, 3, 4, 5, 6, 7) y b) r = 5 bloques, vr= 4 hibridos o variedades comerciales (A B, C, D) y v1 = 13 nuevos tratamientos o nuevas variedades, poblaciones, o hibridos (e, f g, h, I, j, k, l, m, n, o, p, q) (Federer, 1956,1961, 1975).
En ambos ejemplos (a ) y (b) existen r bloques con Nj parcelas en los bloques j=. El proceso de randomización para el diseño de bloques completamente al azar es:
1. Tanto los testigos como los nuevos tratamientos se deben randomizar antes de proceder a distribuir al azar los bloques y las parcelas.
2. Distribuir los testigos al azar en vr + n11 = N1 en el bloque 1. Continuar haciendo el mismo proceso para los bloques restantes (2 a 6) distribuyendolos cada vez al azar en cada bloque. Ejemplo (a) se ha dejado con los testigos solamente para observar la distribución.
3. Distribuir al azar los nuevos tratamientos v1 en las parcelas restantes, lo que equivale a asignar al azar los números o las letras en minúscula a los nuevos tratamientos y asignar las letras o números en orden a las parcelas restantes, (ejemplos a y b).
2. Distribuir los testigos al azar en vr + n11 = N1 en el bloque 1. Continuar haciendo el mismo proceso para los bloques restantes (2 a 6) distribuyendolos cada vez al azar en cada bloque. Ejemplo (a) se ha dejado con los testigos solamente para observar la distribución.
3. Distribuir al azar los nuevos tratamientos v1 en las parcelas restantes, lo que equivale a asignar al azar los números o las letras en minúscula a los nuevos tratamientos y asignar las letras o números en orden a las parcelas restantes, (ejemplos a y b).
Los bloques en el campo deben estar en una area homogenea . El objetivo de los bloques es remover lo mas que se pueda la heterogeneidad en el area experimental y obtener de esta manera las parcelas relativamente homogeneas dentro de cada bloque. Para ver en detalle el analisis tanto para bloques completamente al azar como para lattices se sugiere leer los articulos de Federer, 1956 y 1961.
- Allard R.W. “Principio de la Mejora Genética de las Plantas”. Ediciones Omega, S.A. Barcelona 1967, pp 257-263.
- Clará R. 1980. Informe sobre entrenamiento en el International Crops Research Institute for the Semi-Arid Tropics (ICRISAT—INDIA). Septiembre 24 Diciernbre 6 de 1980. Centro Nacional de Tecnologia Agropecuaria (CENTA). p. 5-6.
- Cochran, W.G., and G.M. Cox. 1992. Experimental Design. John Wiley & Sons, Inc. 2nd ed. Chapters 9 and 10.
- Comstock, R.E., and H.F. Robinson. 1948. The components of genetic variances in popluations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4:254-256.
- D’ Croz-Mason, Nora E. 2004. Maize Breeder, Nebraska University, U.S.A. Referencia personal.
- Doggett, H. And Eberhart, S.A. 1968. Recurrent selection in sorghum. Crop. Sci. 8 (1): 119-121.
- Doggett, H. 1972. The improvement of sorghum in East Africa. 47-59 in Ganga Prasada Rao, N., and House, Leland R. (eds.), Sorghum in Seventies. New Delhi: Oxford & IBH.
- Eckebil, J.P., W.M. Ross, C.O. Gardner, and J.W. Maranville. 1977. Heritability estimates, genetic correlations and predicted gains from S1 progeny tests in three grain sorghum random-mating populations. Crop Sci. 17:373-377.
- Federer, W.T. 1956. Augmented (or Hoonuiaku) designs. Hawaiian Planters= Record, 55, 191-208.
- Federer, W.T. 1961Augmented designs with one-way of elimination of heterogeneity. Biometrics, 17, 447-73.
- Federer, W.T., and D. Raghavarao. 1975. On augmented designs. Biometrics 31, 29-35.
- Fehr,W.R. 1987. Principles of Cultivar Development. Theory and Technique. Vol.1. MacMillan Publishing Company, NY pp 219-246.
- House L.R. 1985. “A guide to Sorghum Breeding”. ICRISAT International Crops Research Institute for the Semi-Arid Tropics, ICRISAT. Patancheru
- P.O., Andhra Pradesh 502 324. pp. 147-148.
- Jan-orn, Jinda, C.O. Gardner, and W.M. Ross. 1976. Quantitative genetic studies of the NP3R random-mating grain sorghum populations. Crop Sci. 16:489-496.
- Obilana, A.T., and M M.El Roubi, 1980. Population improvement and pure line development in sorghum (Sorghum bicolor L. Moench). Cereal Research communications 8 (2), 425-435.
- Lukhele, P. and A.T. Obilana, 1980: S1 selection in a random-mating sorghum population (Sorghum bicolor L. Moench). Presented at the 8th Annual Conference of the Genetics Society of Nigeria, December, 1980.
- Nagur, T. 1981. Cytoplasmic-genetic male sterility and itsutilityin crop improvement. ICRISAT.
- Ross, W.M., and C.O. Gardner. 1983. The mechanics of population improvement in sorghum. In Plant Breeding Methods and Approaches in Sorghum Workshop for Latin America. Sponsored by INTSORMIL, INIA, and ICRISAT, Mexico City. p. 8-38.
- Ross, W.M.. And G.H. Hookstra. 1983. Performance of S1 progenies from a sorghum random-mating population sampled indifferent years. Crop Sci. 23-89-91.
- Ross, W.M., K.D. Kofoid, J.W. Maranville, and R.L. Voigt. 1981. Selecting for grain protein and yield in sorghum random-mating populations. Crop Sci. 21-774-777.
- Russell, W.A. and S.A.Eberthart 1975. Hybrid performance of selected maize lines from reciprocal recurrent and testcross selection programs. Crop Sci. 15:1-4.
- Schutz, W.M., and C.C Cockerham. 1966. The effect of field blocking on gain from selection. Biometrics 22:843-863.
- Stephens, J.C. and R.S. Holland, 1954. Cytoplasmic malesterility for hybrid sorghum seeds production. Agron. J. 46:20-23.
- Verma B. y Lawrence P. 1979. The Improvement of advanced populations..ICRISAT-INDIA, pp 1-11.
- Wall J. S. y Ross W. M. 1975. “ Producción y Usos del Sorgo”. Editorial Hemisferio Sur, Buenos Aires. p. 58.
- Yates, F. 1936. A new method of arranging variety trials involving a large number of varieties. J. Agric. Sci. 26:424-455.
Temas relacionados:
Autores:


Recomendar
Comentar
Compartir

22 de agosto de 2022
Nos brinda la oportunidad de poder formar híbridos comerciales en una planta autógama y también nos emascula para poder hacer mejores cruzamientos en el mejoramiento genético.
Recomendar
Responder

18 de agosto de 2022
Excelente material, sera de gran utilidad
Recomendar
Responder
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Usuarios destacados en Agricultura

Miguel Ángel Taboada  Instituto Nacional de Tecnología Agropecuaria - INTA
Instituto Nacional de Tecnología Agropecuaria - INTA
Investigación, director del Instituto de Suelos del INTA
Estados Unidos de América
Estados Unidos de América

Dr. Ramon León González  North Carolina State University - NCSU
North Carolina State University - NCSU
Investigación / Profesor asociado, biología y ecología de malezas
Estados Unidos de América
Estados Unidos de América




Prof. Luis Ávila  Mississippi State University
Mississippi State University
Profesor Asociado / Investigador
Estados Unidos de América
Estados Unidos de América