Introducción
En su estado natural las pajas utilizadas en la alimentación animal son poco digestibles y, adicionalmente, por efectos asociativos, su inclusión en las raciones afecta negativamente el valor de la energía neta de la dieta. Tales características hacen que la paja sea muy poco utilizada en dietas para ganado de engorda y su uso sea prácticamente inexistente en dietas para ganado lechero, por lo que gran parte de la paja producida es quemada en campo. Sin embargo, las recientes regulaciones ambientales han restringido la quema de pajas y eso ha generado un interés cada vez mayor por darle diversos usos al residuo; entre ellos, aumentar su valor nutrimental con la fi nalidad de utilizarlo en la alimentación del ganado. El problema nutrimental que presentan las pajas es su contenido malo (1%-2%) de carbohidratos solubles combinado con su alto contenido en paredes celulares altamente lignifi cadas.
Adicionalmente, la utilización de la fi bra también está limitada por restricciones a nivel de su organización celular, que hacen al material poco susceptible a hidratarse y, en consecuencia, resistente a las enzimas microbianas. Lo anterior ocasiona bajo índice de reducción del tamaño de la partícula y mayor tiempo de retención ruminal; en consecuencia, el material se acumula en el rumen afectando así el consumo voluntario. La maceración es un proceso mecánico que produce cambios en la integridad estructural y en la densidad de la fracción fi brosa de leguminosas, sufi - cientes para aumentar la digestión de la fi bra y el consumo de materia seca. Por otra parte, la utilización de enzimas fi brolíticas estables en rumen también ha dado resultados positivos en la digestión ruminal de la fi bra y en el consumo de materia seca. Lo anterior supone una posible acción sinérgica cuando se utilizan ambas alternativas en los forrajes de baja calidad.
El objetivo de este estudio fue evaluar la posible interacción de la utilización de una enzima fi brolítica y el proceso de maceración sobre el valor nutrimental de un forraje de baja calidad incluido en un nivel de 40% en la dieta de vacas secas Holstein.
Material y métodos
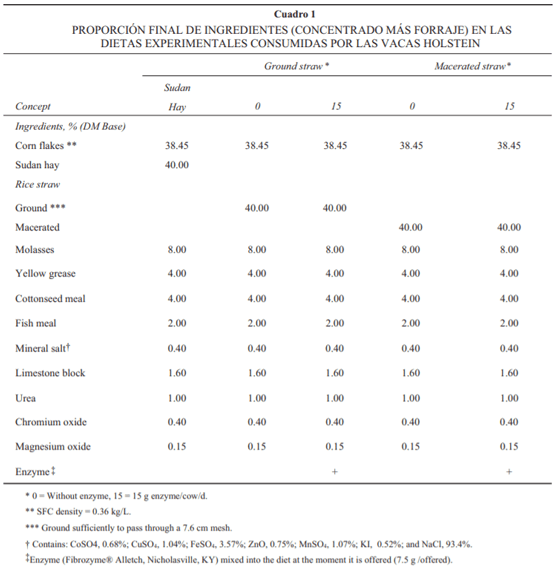
Se utilizaron cinco vacas secas Holstein (633 ± 81 kg) habilitadas con cánulas en rumen y en duodeno (10 cm del esfínter pilórico) en un diseño de cuadrado latino 5 × 5 con el fi n de evaluar la posible interacción entre el uso de una enzima fi brolítica y el proceso de macerado de la paja de arroz sobre las características de digestión y función ruminal. Las cánulas (80 mm di) instaladas en rumen se elaboraron con el material y el procedimiento descrito por Álvarez y Zinn, mientras que las utilizadas para el duodeno fueron cánulas tipo “T” de 25 mm de diámetro interno y se elaboraron con material de tygon inerte.* Las dietas experimentales se presentan en el Cuadro 1 y se formularon combinando un concentrado que contenía 64.07% de maíz en hojuela, 13.33% de melaza, 6.67% de grasa, 6.67% de harinolina, 3.33% de harina de pescado, 0.67% de sal mineralizada, 2.67% de piedra caliza, 1.67% de urea, 0.67% de óxido de cromo y 0.25% de MgO, en proporción de 60:40 con alguno de los siguientes forrajes: a) heno de sudán molido; b) paja de arroz molida; c) paja de arroz macerada; d) paja de arroz molida más enzima (15 g/d); e) paja de arroz macerada más enzima (15 g/d).
La preparación del macerado se realizó pasando la paja por una sola ocasión a través de dos rodillos corrugados (46 × 61 cm) dispuestos horizontalmente uno frente al otro, con abertura de 0.2 mm entre ambos rodillos. Para llevar a cabo la acción de maceración el diferencial de velocidad aplicado entre ambos rodillos fue de 6.8 g (24.5 y 17.7 g) a 62 050 milibares de presión. Una vez obtenido el macerado, éste se molió al igual que el resto de los forrajes en prueba (heno de Sudán y paja de arroz sin macerar) en un molino de forraje de cuchillas,* lo sufi ciente para permitir el paso del material a través de una malla de 7.6 cm. La enzima utilizada fue un preparado comercial,** que es un polvo granular que contiene una combinación de extractos de fermentación de Aspergillus niger y Trichoderma viride y fermentos solubles, protegidos mediante glucosilación, con actividad xilanásica de 100 Uxg-1. La enzima se añadió a la dieta completa (7.5 g/vaca/servida) al momento de ofrecer el alimento. El óxido crómico fue utilizado como marcador inerte para el cálculo, en materia seca, del fl ujo a duodeno y de la excreción fecal. Para efecto de adaptación, 14 días previos al inicio de la prueba, las vacas se asignaron a corraletas individuales de 12.6 m2 con piso de neopreno, comedero individual y bebederos automáticos compartidos. Todas las vacas recibieron el tratamiento 1, siete días antes de iniciar de la prueba. El suministro de las dietas fue ad libitum y se ofreció durante la prueba alimento fresco en dos porciones iguales diariamente a las 0800 y 2000 h. El experimento consistió en cinco periodos experimentales de 14 días (diez para adaptación a la dieta y cuatro para colección de muestras). La recolección de muestras duodenales y fecales se tomaron a cada vaca dos veces al día en los siguientes horarios: día 1, 0650 y 1250 h; día 2, 0800 y 1400 h; día 3, 0950 y 1550 h; y día 4, 1100 y 1700 h. La cantidad de muestra obtenida en cada la recolección fue de aproximadamente 700 mL de quimo duodenal y 400 g de heces (base fresca).
Las muestras de cada vaca se mezclaron en cada periodo de recolección con el propósito de formar una muestra compuesta, que se congeló a –20°C para análisis posteriores. A las 4 h posconsumo (1200 h), en el último día de cada periodo, se determinó el pH*** del contenido ruminal de una muestra obtenida de cada vaca (± 500 mL), mediante una bomba de vacío, a las muestras resultantes se añadieron 2 mL de ácido metafosfórico al 25% (p/v) por cada 8 mL de líquido ruminal previamente fi ltrado a través de tres capas de gasas, posteriormente se centrifugaron (17 000 g durante diez minutos) y el sobrenadante se congeló a –20°C para análisis de ácidos grasos volátiles (AGV).
Una vez determinado el pH ruminal, se procedió a la evacuación total del contenido ruminal en forma manual. El contenido ruminal de cada vaca se mezcló, se pesó y se le tomó una submuestra por triplicado (aproximadamente 1 kg de submuestra total), las cuales se depositaron en refractarios, pesadas y desecadas en estufa a 65°C hasta un peso constante. Posteriormente las muestras se molieron (molino Wiley) lo sufi ciente para el paso a través de una malla de 1 mm y se almacenaron para análisis posteriores. El último día del último periodo experimental se obtuvieron de todas las vacas muestras del fl uido ruminal, éstas se mezclaron y del resultante se obtuvo una alícuota para el aislamiento de bacterias ruminales por centrifugación diferencial. Los procedimientos para el cuidado y el uso de las vacas se realizaron según los procedimientos indicados en el documento de referencias para el cuidado y uso de los animales de experimentación de la Universidad de California. Las muestras generadas se sometieron a todos, o parte de los siguientes análisis: Materia seca (MS; estufa desecando a 105°C hasta obtener un peso constante), cenizas, N Kjeldhal y nitrógeno amoniacal de acuerdo con lo estipulado por la AOAC, energía bruta (EB) (utilizando una bomba calorimétrica adiabática)*, purinas, óxido crómico, almidón, concentración en el fl uido ruminal de AGV y fi bra detergente ácido (FDA) y fi bra detergente neutro (FDN) corregida para cenizas insolubles. La cantidad de materia orgánica microbiana (MOM), así como el nitrógeno microbiano (NM) que fl uyó a duodeno, se calcularon con base en los análisis de las bacterias aisladas en el fl uido ruminal, así como en las muestras obtenidas de duodeno, usando purinas como marcadores microbianos. La materia orgánica fermentada (MOF) en rumen se calculó de acuerdo con la cantidad de MO consumida y las proporciones de MO microbiana y MO total determinadas en duodeno.
El N consumido que escapó de la digestión ruminal (proteína de escape) se consideró como equivalente al total de N que ingresó al duodeno menos la suma de las cantidades de N amoniacal y N microbiano que fl uyeron al duodeno. La tasa de pasaje y digestión ruminal de la fracción de FDN se estimó a través de las variables ruminales determinados para sólidos totales y cantidad de FDN en rumen (kg) y por el fl ujo diario a duodeno de la FDN. La producción de metano en rumen se calculó a partir del balance teórico de fermentación con base en la distribución molar de AGV observada en el fl uido ruminal25 y de la desaparición de MO en rumen. Los datos se analizaron como un diseño de cuadrado latino 5 × 5.26 Los efectos de los tratamientos se contrastaron de la siguiente manera: a) heno de Sudán vs. paja; b) paja molida vs. paja macerada; c) 0 vs. 15 g/d de enzima; y d) interacción de enzima con proceso de macerado. Las medias de los tratamientos se compararon utilizando la prueba de diferencia mínima signifi cativa (DMS).
Resultados
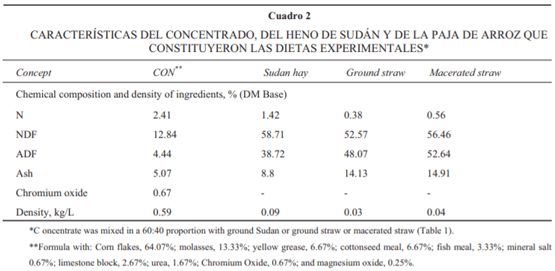
La composición nutrimental y características de densidad del concentrado y de los forrajes utilizados en este estudio se muestran en el Cuadro 2. El contenido de PC del heno de Sudán fue de 8.9%, mientras que la paja de arroz fue de 2.4%. La maceración de la paja de arroz se refl ejó en un incremento del contenido de nitrógeno (47%), de fi bra detergente neutro (7.4%) y de fi bra detergente ácido (9.5%). De igual manera, el macerar la paja aumentó la densidad (p/v) de aquélla en 30%.
Las características del consumo del fl ujo de nutrimentos, así como de la digestión ruminal y del tracto total se muestran en el Cuadro 3. El consumo diario de MS, MO, FDN, FDA, almidón y nitrógeno promediaron 11.97, 10.97, 3.57, 2.62 y 0.204 kg, respectivamente, mientras que el consumo diario de energía bruta promedió 49.1 Mcal, no detectándose diferencia (P > 0.10) signifi cativa entre los tratamientos.
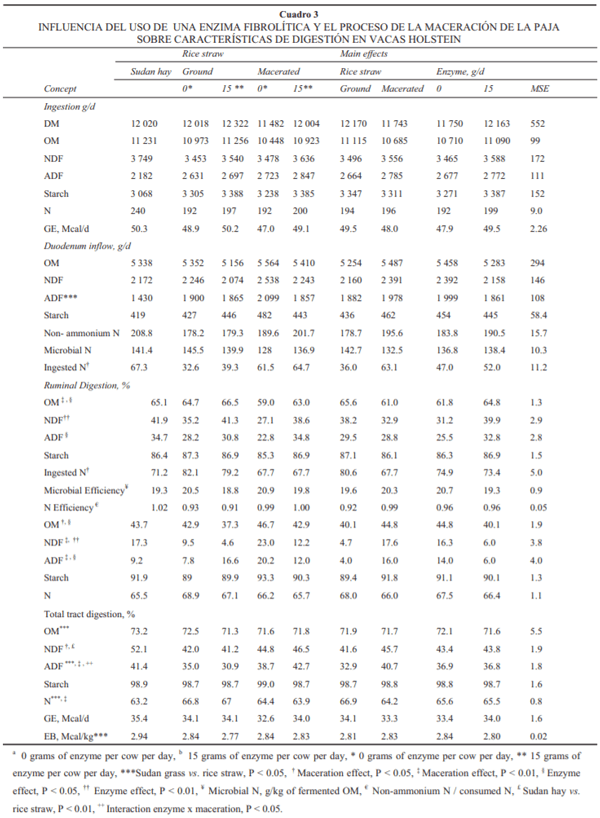
En relación con el fl ujo diario de nutrimentos a duodeno, la dieta con heno de Sudán mostró un menor fl ujo de FDA (P < 0.05) comparado con las dietas que contenían paja, sin diferencias signifi cativas en el resto de las variables evaluadas. No hubo efecto de la maceración o de la adición de la enzima sobre el fl ujo a duodeno de MO, almidón, N no amoniacal y N microbiano. La maceración aumentó el fl ujo a duodeno del N dietético (P < 0.05). Por otra parte, la maceración tendió a incrementar el fl ujo a duodeno de FDN (10.7%, P = 0.09), mientras que la presencia de enzima tendió a disminuirlo (10.8%, P = 0.09).
A nivel ruminal no se detectaron efectos de los tratamientos sobre las variables de efi ciencia microbiana o de efi ciencia de N. La adición de la enzima favoreció un incremento en la digestión a nivel ruminal de la materia orgánica (5%, P < 0.05), de la FDN (21.8%, P < 0.01), y de la FDA (22%, P < 0.05). Con respecto a la digestión ruminal de la FDA, la enzima aumentó (P < 0.05) su digestión en ambos tipos de procesamiento de la paja, pero este efecto tendió a ser más pronunciado (interacción, P = 0.08) cuando la paja fue macerada. Por otra parte, cuando la enzima se añadió a la paja molida resultó en aumento de 19% en la digestión de la FDN en comparación de la paja molida sin enzima, mientras que con la dieta que contenía paja macerada la adición de la enzima se tradujo en un incremento de 42% con respecto a la paja macerada sin enzima.
Contrario al efecto de la enzima, la maceración de la paja de arroz signifi có disminución de la digestión ruminal de la MO (7.0%, P < 0.01), de la FDN (14%, P = 0.06) y del N (16%, P < 0.05); este efecto del macerado sobra la digestión ruminal de la FDN se dio, como se describirá posteriormente, por el marcado aumento de la tasa de pasaje de la FDN en rumen.
A nivel posruminal el heno de Sudán no tuvo efectos sobre las variables evaluadas, mientras que el macerado incrementó la digestión de la MO (P < 0.05), la FDN (P < 0.01), la FDA (P < 0.01) y tendió a disminuir la digestión aparente del N (P = 0.06). La enzima mostró un efecto contrario al macerado en este sitio, ya que disminuyó la digestión de MO (P < 0.05), FDA (P < 0.05), FDN (P < 0.01) sin efecto alguno en las otras variables evaluadas.
En el tracto total se observó una digestión mayor (P < 0.05) para la MO, FDN y energía bruta (3%, 19% y 4%, respectivamente) para la dieta que contenía heno de Sudán molido con respecto a las dietas que contenían paja de arroz; lo anterior se refl ejó en un mayor contenido de la energía digestible (Mcal/kg) de la dieta para la ración con heno de Sudán (4%, P < 0.01).
En general, la maceración se refl ejó en una disminución (4.0%, P < 0.01) de la digestibilidad aparente del N dietético y en un incremento neto (P < 0.01) de la digestión en tracto total de la FDA.
Cuando la paja fue macerada, la digestibilidad de la FDA aumentó en 24%, pero con la presencia de la enzima el incremento en la digestión resultó en 38% (interacción, P < 0.05). Por otra parte, no hubo efecto (P > 0.10) de adición de la enzima sobre las variables evaluadas en tracto total.
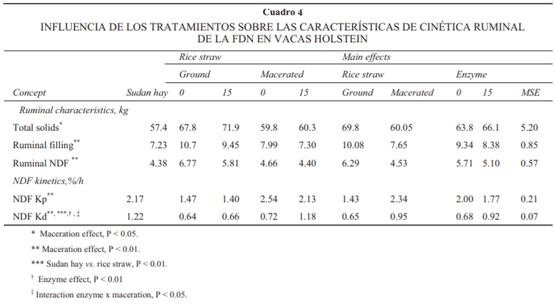
Los efectos de los tratamientos sobre la cinética ruminal de la fracción de la FDN se muestran en el Cuadro 4. Comparado con la dieta que contenía paja, la dieta con heno de Sudán resultó en mayor tasa de digestión (33%, P < 0.01) y tendió a presentar una menor cantidad de FDN en rumen (19%, P = 0.09). Hubo efecto de interacción enzima × macerado sobre la tasa de digestión ruminal de FDN. La presencia de la enzima en la dieta con paja molida tuvo efecto nulo sobre esta variable (0.66% vs. 0.64%/h, con y sin enzima, respectivamente), mas cuando la enzima se combinó con la paja macerada aumentó en 78% la tasa de digestión de la FDN (P < 0.05) respecto de la paja molida. La maceración de la paja disminuyó el contenido de sólidos (13%, P < 0.05) y de la cantidad de FDN en rumen (28%, P < 0.01) e incrementó la tasa de pasaje y la tasa de digestión ruminal de la FDN en 26% y 52%, respectivamente (P < 0.01). Aun cuando la presencia de la enzima no infl uyó (P > 0.10) a nivel ruminal en el contenido de fi bra detergente neutro, cantidad de sólidos o tasa de pasaje de la FDN, promovió un incremento de la tasa de digestión de la FDN (43%, P < 0.01).
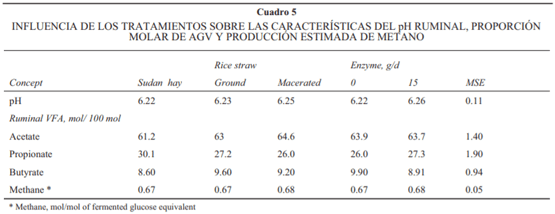
El efecto de los tratamientos sobre el pH ruminal y el perfi l de AGV se muestran en el Cuadro 5. La dieta con heno de Sudán tendió a mostrar una menor (P = 0.09) proporción de acetato comparado con las dietas con paja, sin diferencia en el resto de las variables evaluadas. Por otra parte, no hubo efecto por el uso de la enzima o por el tipo de procesado de la paja sobre el pH ruminal, los perfi les de AGV o la producción de metano.
Discusión
La gran diferencia en el contenido de N de la paja macerada con respecto a la paja molida pudo ser resultado del procedimiento de muestreo. Aun así, cambios en la composición de la fracción nitrogenada y fracción fi brosa después de que el material fue sometido al proceso de maceración se han observado en leguminosas como resultado directo de la pérdida de hojas, y por la pérdida de material soluble de las células durante el procesado y el manejo del material. Como en el presente estudio, Petit et al. detectaron un incremento del 17% del contenido del N posterior a la maceración del heno de pasto Timothy; estos investigadores explican que la insolubilidad de la fracción nitrogenada se ve disminuida por el macerado, por lo que a mayor madurez de la planta se espera un cambio mayor en el contenido de esta fracción. Por otra parte, las modifi - caciones en la estructura de la pared celular y tamaño de la partícula resultan en cambios de algunas propiedades físicas, entre ellas, la densidad. El aumento en la densidad (g/L), como resultado de la maceración, es consistente con resultados obtenidos en estudios previos.
Las vacas utilizadas en este estudio fueron vacas secas y consumieron, en promedio, 4% menos que lo esperado, de acuerdo con las predicciones del NRC; esta ligera baja en el consumo es consecuencia del medio ambiente en que se desarrolló el experimento (corraletas individuales) y de la habilitación de los animales con cánulas en rumen y duodeno. Los efectos debidos a la adición de enzimas exógenas frecuentemente están más relacionados con la digestibilidad de la FDN que con el consumo de la materia seca. Aunque algunos estudios han informado la ausencia del efecto de la enzima sobre el comportamiento de consumo, esto no siempre sucede. De igual forma, mientras que en algunos estudios el consumo de MS se aumenta después de que el forraje ha sido macerado, en otros no se ha observado este efecto. Las diferencias en las respuestas al consumo de dietas que contienen forrajes a los que se le han añadido enzimas fi brolíticas, o que han sido macerados, pueden ser consecuencia de diversos factores, como el tratamiento previo y el nivel de forraje en la dieta, la duración del periodo de prueba, el estado productivo del animal, la cantidad de enzima utilizada y, en el caso del macerado, la intensidad del proceso de maceración. En el caso particular del macerado, en la literatura revisada no se indica con precisión la intensidad del proceso o las características físicas (densidad) del producto macerado. Estos últimos factores pueden ser relevantes en cuanto al comportamiento de consumo cuando es incluido a las dietas este tipo de material.
La enzima y el macerado tendieron (P = 0.08) a mostrar sinergia en la digestión ruminal de la FDA; sin embargo, estas diferencias no se detectaron para la digestión ruminal de la FDN aunque numéricamente la diferencia fue grande (42%) para la combinación enzima × macerado comparado con la combinación enzima × molido. La hipótesis aquí planteada se fundamentó sobre la base de que el macerado promueve cambios en la estructura (organización) de las paredes celulares que le confi eren propiedades físicas ventajosas para su digestión, las cuales permiten mejor hidratación y mayor exposición a las enzimas microbianas, lo que, aunado a la presencia de la enzima exógena, pudiese tener un efecto sinérgico importante sobre la digestión de la fibra.
Con respecto a lo anterior, en un estudio previo se comparó in vitro el efecto de la utilización de una enzima exógena pura (mezclada 100% con saliva McDougall) o combinada con fl uido ruminal (80% saliva de McDougall y 20% de fl uido ruminal); la combinación de la enzima exógena con el fl uido ruminal promovió importantes aumentos en la tasa de desaparición neta (2.7% vs. 24.2%) de esa fracción. Esta sinergia se observó también en el presente estudio, ya que hubo un aumento (1.18% vs. 0.72%; interacción, P < 0.05) de la tasa de digestión ruminal (kd) de la FDN cuando la enzima se combinó con la paja macerada. Este resultado adquiere relevancia principalmente en los sistemas de alimentación en los que esté comprometida la actividad celulolítica; por ejemplo, dietas altas en concentrados que resulte en pH ruminal bajo, o bien en dietas con alto contenido de forraje donde la cantidad de FDN limite el consumo total de energía; por ejemplo, dietas para ganado en crecimiento o bien dietas destinadas para ganado lechero.
El resultado más consistente cuando se utilizan enzimas fi brolíticas exógenas es el aumento en la digestión ruminal de la fi bra sin cambios en la digestión de esa fracción a nivel de tracto total, mientras que el efecto de la enzima sobre la digestión ruminal de la materia orgánica invariablemente es un refl ejo directo de los cambios de la digestión ruminal de la fracción de fi bra; por consiguiente, las diferencias de mayor magnitud en la digestión ruminal de la fi bra hacen posible que las diferencias en la digestión ruminal de la MO sean mayores. De la misma manera, los cambios en la digestión ruminal de la fi bra aparentemente están supeditados a la cantidad de FDN presente en la dieta. Por ejemplo, se han observado aumentos similares (22% vs. 18%) en la digestión ruminal de la FDN en dietas con nivel de forraje similar e igual tipo y cantidad de enzima que el utilizado en el presente estudio.
Por otra parte, Zinn y Salinas observaron 28% de aumento en la digestión ruminal sin cambios en la digestión de la fi bra detergente neutro a nivel de tracto total cuando se usó este mismo tipo de enzima en novillos con dietas de fi nalización (< 12% de forraje); por lo anterior, debe considerarse que la digestión de la FDN con ese tipo de dietas generalmente está deprimida por el pH ruminal que prevalece, y el uso de enzimas en esas condiciones puede refl ejar resultados de mayor magnitud comparado con el grupo testigo. En estudios conducidos con dietas altas en forraje (>60%), la utilización de enzimas fi brolíticas también ha resultado en incrementos en la digestión de la FDN tanto en nivel ruminal como en tracto total; en contraste, existen estudios en que se utilizaron niveles altos de forraje y no se observaron diferencias en la digestión ruminal de la FDN, pero sí aumentos en la digestión de esa fracción a nivel de tracto total. Sea como fuere, las enzimas exógenas aparentemente no infl uyen en el contenido de FDN, sólidos en rumen o en la tasa de pasaje de la fi bra detergente neutro, por lo que su efecto principal representa un incremento de la tasa de digestión de la FDN tanto in vitro como in vivo.
Ello explica la menor cantidad de FDN fl uyendo a duodeno detectada en el presente estudio. En estudios en que se ha evaluado el proceso de maceración en la alfalfa, se han observado aumentos en la digestibilidad ruminal y en tracto total de la MO, FDN y FDA, sin efectos sobre la digestibilidad del N dietético. Sin embargo, Torrentera et al. no observaron diferencias en la digestión ruminal de la MO y de la FDN cuando compararon paja de arroz macerada con paja de arroz molida, que se incluyó en 20% en dietas para ganado de engorda. Como se mencionó en la sección de resultados, el macerado afectó negativamente la digestión ruminal de la MO, N y FDN. Tal resultado es congruente con lo observado en el presente estudio en relación con el incremento de fl ujo a duodeno de las fracciones de FDN, FDA y N, así como de las variables de cinética ruminal, ya que el macerado resultó en aumento de la tasa de pasaje ruminal de la FDN (52%; P < 0.01, Cuadro 4), que disminuye el tiempo de retención en rumen y la digestión ruminal aparente. Durante la evacuación ruminal se notó, para el caso de la paja macerada, que el contenido ruminal se presentaba relativamente homogéneo, mientras que con la paja molida presentaba cierta estratifi cación de las partículas. En un estudio previo, el nivel de estratifi cación del contenido ruminal en dietas que contenían paja se relacionó con la disminución de la tasa de pasaje; en tanto que, en un estudio realizado con leguminosas, en que se detectó aumento en la digestibilidad ruminal de la MO y de la FDN, el tiempo de retención ruminal no se afectó por la maceración.
Según resultados de estudios previos, la maceración incrementó en tracto total la digestión de la FDN y de la FDA como resultado de mayor digestión posruminal de esas fracciones. El efecto sobre la disminución de la digestibilidad ruminal del N debido al macerado no es claro, la mayoría de los estudios no informan efectos negativos signifi cativos sobre esa variable.
El macerado de la alfalfa en presentación heno o en forma de ensilado no ha resultado en cambios de la tasa de pasaje ruminal. Los estudios indican que la maceración cambia la estructura de la partícula de tal forma que permite mayor hidratación, con más rápida reducción de su tamaño e importante cambio de la gravedad específi ca, todos esos factores promueven aumento en la tasa de pasaje. Sin embargo, esto último puede variar, ya que la relación entre el cambio de las características físicas de las partículas durante su estancia en rumen, respecto de la tasa de pasaje y de digestión, varía por el tipo de material y las características fi sicoquímicas de los tejidos vasculares de la planta. Lo anterior indica la posibilidad de que los forrajes de baja calidad respondan de manera distinta al proceso de macerado. Es bueno realizar más estudios para seguir evaluando las respuestas al macerado de forraje de baja calidad, como las pajas.
Una mayor digestión ruminal de la FDN y una menor digestión ruminal del N de la dieta ha sido resultado común cuando se han comparado dietas que contienen heno de Sudán con las que contienen pajas, y está supeditado principalmente a diferencias en las propiedades fi sicoquímicas de ambas fracciones entre las fuentes de forraje. La tasa de pasaje y la tasa de digestión de la FDN de los henos de gramíneas generalmente es mayor que el de las pajas. Aun cuando en el presente estudio la tasa de digestión fue mayor (1.22% vs. 0.80%/h; P < 0.01) para el heno de Sudán respecto de las dietas que contenían paja de arroz, la maceración de la paja incrementó la tasa de digestión de la FDN a un valor similar al del heno Sudán (1.18% vs. 1.22 %/h). De la misma forma, cuando se comparó el valor promedio de la tasa de pasaje de la FDN de la paja molida (1.44 %/h) con la tasa de pasaje del heno de Sudán (2.17%/h, Cuadro 4) se registró en una diferencia signifi cativa (P < 0.05); sin embargo, el aumento en la tasa de pasaje debido al efecto de maceración (2.34%/h) no permite detectar diferencias signifi cativas (P > 0.18) cuando se compara el heno de Sudán con el valor promedio de los tratamientos de paja. Asimismo, se observan valores más altos en la digestibilidad de la MO, FDN y EB de la dieta a nivel de tracto total cuando se comparan henos de gramíneas con dietas que contienen pajas.
En promedio, el pH ruminal observado fue de 6. y concuerda con otros estudios en que se usó la proporción de concentrado: forraje similar a la aquí descrita. En estudios previos en dietas con bajo, mediano o alto contenido de forraje, el uso de enzimas fi brolíticas no resultó en cambios del pH ruminal, perfi les de AGV o la producción estimada de metano. De igual forma, la maceración no ha mostrado efectos sobre estas variables ruminales.
La enzima fi brolítica aquí probada actuó en forma sinérgica con el proceso de maceración, ello resultó en una mejora del valor nutrimental de forrajes de baja calidad. Se espera que los efectos benéfi cos por el uso de las enzimas fi brolíticas sobre la digestión ruminal se aumenten a medida que se disminuya el tiempo de retención ruminal de los componentes fi brosos de la dieta.