Introducción
La alta concentración salina de los suelos es uno de los principales factores que restringe el crecimiento y productividad de los cultivos (Cruz et al. 2003); por lo tanto, comprender la respuesta de las plantas bajo condiciones de estrés es importante para proponer soluciones efectivas al problema de la salinidad (Larcher 2003).
La característica general de los suelos salinos es la presencia de altas concentraciones de sales solubles, lo cual incrementa el potencial osmótico de la solución del suelo, causando estrés fisiológico (Wahome et al. 2001). Este problema es común en las regiones áridas y semiáridas, como una consecuencia del déficit hídrico, por la escasez de precipitaciones y la alta tasa de evaporación (Ferreira et al. 2001).
La salinidad origina reducción del crecimiento de los cultivos al afectar negativamente la germinación y/o la capacidad de emerger de las plántulas. Igualmente retarda el crecimiento de las plantas a través de su influencia sobre varios procesos fisiológicos, tales como: fotosíntesis, conductancia estomática, ajuste osmótico, absorción de iones, síntesis de proteínas, síntesis de ácidos nucleicos, actividad enzimática y balance hormonal (Mizrahi et al. 1988; Bethke & Drew 1992; Hakim & Rhaman 1992), además puede afectar el proceso de transporte de agua e iones, lo que promueve toxicidad iónica y desbalance nutricional (Lerner 1985; Chartzoulakis & Klapaki 2000). En consecuencia, las variables de crecimiento vegetativo tales como: masa seca, altura de la planta y área foliar, entre otras, son severamente afectadas por la presencia de sales (Rush & Epstein 1981; Alarcón et al. 1993).
Los cultivos responden de manera particular a la salinidad, algunos producen rendimientos aceptables a altas concentraciones de salinidad, mientras que otros son sensibles a bajas concentraciones (Serrano 1996). Tester & Davenport (2003) señalaron que la intensidad con que cada condición de estrés afecta el crecimiento y desarrollo de las plantas depende de varios factores entre los cuales se incluye: la especie, el cultivar, estado fenológico, tipos de sales solubles, intensidad y duración del estrés y condiciones edafoclimáticas. Se ha reportado que la salinidad reduce la tasa de crecimiento y, consecuentemente, la producción de los cultivos a través de la disminución de la eficiencia fotosintética, ya sea por disminución de la asimilación de fotosintetizado, posiblemente por la reducción del conjunto de nucleótidos y el gasto adicional de energía, por declinación de la conductancia estomática o por altos niveles de los iones sodio y cloro en el tejido foliar (Bethke & Drew 1992; Nieman et al. 1998; Chartzoulakis & Klapaki 2000). Por lo tanto, el efecto osmótico no sólo causa un simple efecto físico sobre la reducción de la presión de turgor de las células de la planta, sino que involucra alteraciones bioquímicas o fisiológicas que envuelven la expresión de los genes (Termaat et al. 1985).
Además de los ajustes fisiológicos que se producen en las plantas para tolerar la salinidad, éstas sufren cambios morfológicos y anatómicos, lo que modifica la coordinación entre diferentes órganos o tejidos, incrementa la habilidad para capturar o conservar recursos o aumenta su tolerancia al factor causante del estrés, como lo es la modificación de la densidad o el índice estomático, que ocurren frente a algunos tipos de estrés (Salas et al. 2001; Kakani et al. 2003; Neogy et al. 2003; Benavides-Mendoza et al. 2004). La salinidad induce variaciones en el índice y densidad estomática del tejido foliar en especies como Capsicumannuum L. y Lycopersicum esculentum Mill (Bethke & Drew1992; Salas et al. 2001).
Según Martín et al. (1994), ésta también se relaciona con cambios fisiológicos y anatómicos a nivel foliar, lo que puede ocasionar modificaciones en la frecuencia estomática de las hojas, en el espesor de la cutícula y alteraciones en la resistencia estomática al intercambio gaseoso, entre otros (Kebede et al. 1994).
Aparentemente, los cambios anatómicos son respuestas morfogenéticas de la planta para contrarrestar los efectos negativos de las sales, cambios que pueden ser importantes en la eficiencia del uso del agua y la tolerancia de la planta al estrés salino (Shannon 1985; Pio et al. 2001). En consecuencia, el conocimiento de la respuesta de una especie vegetal cultivada bajo condiciones de estrés salino es importante para poder establecer técnicas alternativas de manejo con la finalidad de aminorar los efectos perjudiciales de las sales (Ferreira et al. 2001).
El objetivo del presente trabajo fue evaluar el efecto de la salinidad sobre el espesor de la lámina foliar y la densidad e índice estomático de las láminas foliares de plantas de lechosa (Carica papaya) al inicio de la etapa reproductiva.
Materiales y Métodos
El ensayo se llevó a cabo en las zonas de cultivo del Decanato de Agronomía de la Universidad Centroccidental Lisandro Alvarado, municipio Palavecino, Cabudare, estado Lara. El material vegetal utilizado en esta investigación se obtuvo a partir de semillas certificadas de lechosa (Carica papaya) cv Maradol, procedentes de México y producidas por la empresa Semillas del Caribe.
Previo a la siembra, las semillas fueron sometidas a un tratamiento pregerminativo por 24 h con agua destilada, luego se sembraron a razón de 8 semillas/contenedor.
El sustrato estuvo constituido por tierra:arena:humus de lombriz en proporción 2:1:1, y fue químicamente desinfectado con Dazomet a razón 45 g m-3 de sustrato. Las plántulas se protegieron inicialmente del sol cubriéndolas con restos de pasto, el cual fue retirado progresivamente. Posterior a la emergencia se realizó un entresaque, dejando las tres plantas más vigorosas.
Los tratamientos evaluados fueron cuatro concentraciones de salinidad ajustados a conductividades eléctricas (CE) de 0,001; 2; 4 y 8 dS m-1. Los tratamientos salinos se comenzaron a aplicar a los siete días después de la emergencia, con una frecuencia de aplicación interdiaria. Previo a ellos, las plántulas fueron regadas con agua destilada.
Los niveles de salinidad se prepararon con cloruro de sodio (NaCl) grado analítico (Makhija & Jindal 1983). Para calcular la cantidad de NaCl necesaria para cada tratamiento se estableció una curva de calibración que relacionó cada uno de los tratamientos de salinidad con la cantidad (g) de NaCl disueltos en un litro de agua destilada; la ecuación de regresión calculada fue Y = 4,5183x – 4,3444, r = 0,9995, r2 = 0,9991.
Diseño de experimento
El estudio estuvo regido estadísticamente bajo un diseño de experimento enbloques al azar, con cuatro tratamientos y cuatro repeticiones, en el que se consideraron como tratamientos a los niveles de salinidad medidos por la conductividad eléctrica. La unidad experimental estuvo conformada por tres contenedores, cada uno con tres plantas, para un total de 36 plantas por tratamiento y 144 en total.
Al inicio de la etapa reproductiva se colectaron, de cada unidad experimental, dos hojas de plantas distintas, para un total de ocho láminas foliares por tratamiento, las mismas se seleccionaron del nudo número seis en sentido basípeto. De cada lámina se fijó en FAA 70% (Formaldehído:Ácido acético:alcohol) (Cutler 1978) el tercio medio de la lámina y posteriormente se sometieron a aclarado segmentos de aproximadamente 1 cm2, utilizando para ello una solución de hipoclorito de sodio al 40% por 24 h a temperatura ambiente. La tinción se realizó con safranina y se hicieron montajes semipermanentes con agua-glicerina (1:1) (Salas et al. 2001).
Los valores promedios de la densidad (DE) y el índice estomático (IE) se determinaron en los aclarados epidérmicos y en campos de 400×. El espesor de la lámina se determinó en secciones transversales a mano alzada y se midió con la ayuda de un microscopio óptico Olympus BX40 en un campo de 400× y provisto de una escala.
Se realizaron de 128 observaciones correspondientes a las cuatro repeticiones por cada tratamiento. El IE se calculó utilizando la ecuación sugerida por Wilkinson (1979):
NE × 100
IE = –––––––––––––
NCE + NE
en la cual, NE es el número de estomas por campo de observación y NCE es el número de células epidérmicas típicas en el campo de observación. La densidad estomática se obtuvo contando el número de estomas en un área de 4,347 mm2, equivalente al diámetro del campo observado, con aumento de 400×. Se realizó el análisis de la varianza para las variables índice y densidad estomática y espesor de la lámina, usando el paquete estadístico Statistics (2000), previa verificación de las hipótesis del modelo. Al detectarse diferencias significativas entre los tratamientos, se realizaron pruebas de comparación de medias de acuerdo a Tukey, con un nivel de 5% de probabilidad.
Resultados y Discusión
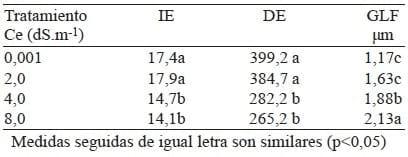
Los resultados demostraron que la salinidad del agua de riego promovió variaciones en el índice estomático (IE) y la densidad estomática (DE) y en el espesor de los tejidos foliares. El IE varió en un rango de 14,1 a 17,9 y su tendencia fue disminuir a consecuencia del incremento de las concentraciones de sal aplicadas.
Así, el material vegetal analizado quedó clasificado en dos grupos estadísticos (Tabla 1). A pesar de que Metcalfe & Chalk (1979) mencionaron que el IE es unacaracterística de valor diagnóstico muy utilizada en sistemática de plantas, por mantenerse sin alteraciones,Wilkinson (1979) y Kürschner et al. (1998) señalaron que es una variable afectada por condiciones estresantes tanto ambientales como nutricionales. Bethke & Drew (1992) y Salas et al. (2001) encontraron que esta variable se modificó en plantas de pimentón y tomate, respectivamente, al ser sometidas a estrés salino.
El incremento de la salinidad disminuyó el número de estomas en la superficie abaxial de las láminas foliares.
La prueba de Tukey permitió clasificar el material vegetal analizado, de acuerdo a la DE, en dos grupos estadísticos (Tabla 1), valores que concuerdan con lo establecido para el envés de las hojas de las plantas con metabolismo C3 (Leegod 1993).
Tabla 1. Efecto de la conductividad eléctrica sobre el índice estomático (IE), la densidad estomática (DE) y el grosor de la lámina foliar (GLF) de plantas de Caricapapaya L.
Estos resultados coinciden con los obtenidos por Benavides et al. (2004) en ocho genotipos de maíz, ya que los valores de la DE de las hojas de este cultivo disminuyeron en la superficie abaxial, al someterlas a estrés salino y a juicio de este mismo autor, este ajuste en la estructura epidérmica pudiese estar relacionado con la eficiencia en el uso del agua en el sentido de que incrementan su habilidad de capturar o conservarla; a su vez contrastan con los obtenidos por Willadino et al. (1999) quienes encontraron que la salinidad no promovió alteraciones en el número de estomas a nivel foliar en plantas de maíz. Sin embargo, Salas et al. (2001) reportaron que el incremento de la salinidad redujo la DE en la superficie adaxial de las hojas de tomate.
La presencia de sales afectó el grosor de las láminas foliares (Tabla 1). Al comparar las medias entre los tratamientos, se encontró que las plantas regadas con agua a 0,001 y 2 dS.m-1 de conductividad eléctrica presentaron un comportamiento estadísticamente similar entre sí e inferior (1,4 mm en promedio) al de los tratamientos regados con agua a 4 dS.m-1 y 8dS.m-1 (1,88 y 2,13 mm, respectivamente), resultados que coinciden con los obtenidos por Kebede et al. (1994) en plantas de tomate, donde el grosor de la lámina incrementó, como consecuencia de un aumento del grosor de la cutícula, al ser sometidas a estrés salino.
Aparentemente, estos cambios anatómicos a nivel foliar en las plantas de lechosa cv Maradol constituyen una respuesta morfogenética que contrarresta losefectos negativos de las sales; los mismos pueden ser importantes en la eficiencia del uso del agua y la tolerancia de la planta al estrés salino (Shannon 1985; Pio etal. 2001) por cuanto es conocido que la salinidad del suelo limita la disponibilidad del agua para las plantas debido al efecto osmótico que se produce (James et al. 1982). En este sentido, la reducción de la DE adaxial parece indicar una respuesta de la planta para aumentar su resistencia estomática promedio y evitar el exceso de transpiración (Rubino et al. 1989; Salas et al. 2001).
Conclusión
La salinidad del agua de riego causó una reducción del índice estomático y del número de estomas por unidad de área al inicio de la etapa reproductiva. Por el contrario, el espesor del tejido foliar se incrementó por efecto de los tratamientos.
Bibliografía
Alarcón, J., M. Sánchez-Blanco, M. Bolarin & A. Torrecillas. 1993. Water relations and osmotic adjustments in Lycopersicum esculentum and L.pennelli during short-term salt exposure and recovery. Physiol. Pl. 89: 441-447.
Benavides, A., R. Dávila, F. Rincón, H. Ramírez & L. Fuentes. 2004. Respuesta de la densidad estomática y de células tabulares de maíces bajo estrés por NaCl. http://www.uaaan.mx/DirInv/Resul_PI04/MEMORIA_20 04/IngAgricola/ABenavidesMendoza-2.doc.182-186 p.
Bethke, P. & M. Drew. 1992. Stomatal and nonstomatal components to inhibition and photosynthesis in leaves of Capsicum annum L. during progressive exposure to NaCl salinity. Pl. Physiol. 99: 219-226.
Chartzoulakis, K. & G. Klapaki. 2000. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hort. 86(3): 247-260.
Cruz, J., C. Pelacani, S. Filho & W. Dos Santos. 2003. Production and partitioning of dry matter and stomatal conductance of Rangpur lemon under salt stress. Rev. Bras. Frutic. 25(3): 528-531.
Cutler, D. 1978. Plant Anatomy. Experiment and interpretation. Part I. Cells and tissues. Second edition. Edward Arnold, London.
Ferreira, R., F. Tavora & F. Ferreyra. 2001. Dry matter partitioning and mineral composition of roots, stems and leaves of guava grown under salt stress conditions. Pesq. Agropecu. Bras. 36(1): 79-88.
Hakim, M & L. Rhaman. 1992. Tolerance of some tomato cultivars to salinity. Acta Hort. 323: 183 189.
James, D., R. Hansks & J. Jurinak. 1982. Modern irrigated soils. Wiley. New York.
Kakani,V., K. Reddy, D. Zhao & A. Mohammed. 2003. Effects of ultraviolet-Bradiation on cotton (Gossypium hirsutum L.) morphology and anatomy. Ann. Bot. 91: 817-826.
Kebede, H., B. Martín, J. Nienhuis & G. Jung. 1994. Leaf anatomy of two Lycopersicon species with contrasting gas exchange properties. Crop Sci.34: 108-113.
Kürschner,W., I. Stulen, F. Wagner & P. Kuiper. 1998. Comparison of palaeobotanical observations with experimental data on the leaf anatomy of durmast oak [Quercus petraea (Fagaceae)] in response to environmental change. Ann. Bot. 81: 657-664
Larcher,W. 2003. Physiological plant ecology. Fourth edition. Springer-Verlag. Berlin.
Leegod, R. 1993. Carbon dioxide concentrating mechanisms. In: Plant biochemistry and molecular biology (Lea, P. J. & R.C. Leegod, eds.). JohnWiley & Sons, Ltd. Chichester.
Lerner, H. 1985. Adaptation to salinity at the plant cell level. Plant and Soil. 89: 3-14.
Makhija, M. & P. Jindal. 1983. Effect of different soil salinity leves on seed germination and seedling growth in papaya (Carica papaya L.). SeedRes. 11: 125-128.
Martín, B., H. Kebede & C. Rilling. 1994. Photosynthetic differences among Lycopersycum species and Triticun aestivum cultivars. Crop Sci. 34: 113-118.
Metcalfe, C. & L. Chalk. 1979. Anatomy of the dicotyledons. Vol. 1. Oxford Clarendon Press, London.
Mizrahi, Y., E. Taleisnik, V. Kagan-zur, Y. Zohar, R. Offenbach, E. Matan & R. Golan. 1988. A saline irrigation regime for improving tomato fruit quality without reducing yield. J. Amer. Soc. Hort. Sci. 113: 202-205.
Neogy, M., J. Datta, A. Roy & S. Mukerji. 2003. Studies on phytotoxic effect of aluminum of growth and some morphological parameters of Vignaradiata L. Wilczek. J. Environ. Biol. 23: 411 416.
Nieman, R., R. Clark, D. Pap, G. Ogata & E. Maas. 1998. Effects of salt stress on adenin and uridini nucleotide poll. J. Exp. Bot. 39: 301-309.
Pio A., C. Horst, H. Martínez, C. Martínez & P. Mosquim. 2001. Características fisiológicas de porta-enxertos de videira em solução salina. Sci.Agric. 58(1): 139-143.
Rubino, P., E. Tarantino & F. Rega. 1989. Relationship between soil water status and stomatal resistance of tomatoes. Irrigazione e Drenaggio 36: 95-98.
Rush,W. & E. Epstein. 1981. Comparative studies on the sodium, potassium and chloride relations of a wild halophytic and a domestic salt-sensitive tomato species. Pl. Physiol. 68: 1308-1303.
Salas, J., M.E. Sanabria & R. Pire. 2001. Variación en el índice y la densidad estomática en plantas de tomate (Lycopersicum esculentum Mill.) sometidas a tratamientos salinos. Bioagro 13(3): 99-104.
Serrano, B. 1996. Efecto de la salinidad sobre el crecimiento y desarrollo de algunos cultivares y portainjertos de vid. Tesis de Maestría. Decanato de Agronomía. Universidad Centroccidental Lisandro Alvarado. Cabudare. Venezuela.
Shannon, M. 1985. Principles and strategies in breeding for higher salt tolerance. Plant and Soil 89: 227-241.
Statistic. 2000. Analytical software. V.7,0. Florida.
Termaat, A., J. Passioura & R. Munns. 1985. Shoot turgor does not limit shoot growth of NaCl-affected wheat and barley. Pl. Physiol. 77: 869-872.
Tester,M. & R. Davenport. 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 91: 503-527.
Wahome, P., H. Jesch & I. Grittner. 2001. Mechanisms of salt stress tolerance in two rose rootstocks: Rosa Chinensis “Major” and R. rubiginosa. Sci.Hort. 87: 207-216.
Wilkinson, H. 1979. The plant surface (mainly leaf) In: Anatomy of dicotyledons (Metcalfe, C.R. & L. Chalk, eds.). Second edition. Vol. 1. Oxford ClaredonPress. London.
Willadino, L., M. Martins, T. Camara, A. Goncalves & G. Dias. 1999. Resposta de genótipos de milho ao estresse salino em condições hidropônicas. Sci. Agric. 56(4): 1209-1213.