Efecto del zinc en la alimentación porcina
Publicado: 11 de noviembre de 2020
Por: Alberto Quiles. Departamento de Producción Animal. Facultad de Veterinaria. Universidad de Murcia. España
INTRODUCCIÓN
La Producción Porcina es cada vez más exigente y tecnificada como consecuencia de los avances en Reproducción Porcina, en Mejora Genética o en el manejo de los animales, lo que está ocasionando que tengamos que prestarle una mayor atención a los requerimientos de ciertos oligoelementos, para poder llevar a cabo el crecimiento tisular esperado. Si a la mejora de los índices productivos, añadimos que ha disminuido la ingesta voluntaria de los cerdos, podemos entender que las recomendaciones efectuadas por los expertos tengan que ser revisadas periódicamente para adaptarse a las necesidades reales de los oligoelementos en las condiciones actuales; mejorando, a la vez, su biodisponibilidad.
Ahora bien, hemos de decir que al amparo de estas últimas reflexiones, en ocasiones se ofrecen a los cerdos niveles de oligoelementos muy superiores a las recomendaciones (Tabla 1), aportándose en ocasiones 3 ó 4 veces más de lo necesario. Si bien ello, en la mayoría de las ocasiones, no tiene consecuencias negativas de toxicidad para los animales, sí tiene repercusiones ambientales, sobre todo en el caso del cobre y del zinc, ya que su excreción contribuye a la contaminación de los suelos y aguas, pudiendo ser tóxico para la microflora del suelo. De ahí que el Comité Científico para la Nutrición Animal (SCAN) de la Unión Europea propuso disminuir los niveles máximos de zinc, lo que posteriormente quedó plasmado en el Reglamento (CE) nº 1334/2003 por el que se modifican las condiciones para la autorización de una serie de aditivos en la alimentación animal pertenecientes al grupo de oligoelementos, en el cual el límite máximo del zinc queda establecido en 150 ppm para la especie porcina.
La carencia de determinados oligoelementos (sustancias minerales necesarias en pequeñas cantidades para el normal funcionamiento del organismo) puede constituir un factor limitante para la Porcinotecnia, concretamente en el caso que nos ocupa el zinc tiene un efecto directo sobre el crecimiento post-destete de los lechones y sobre las características seminales del verraco.
Antes de analizar estos aspectos, entendemos que el lector debería tener unos conocimientos generales sobre capítulos tales como: funciones fisiológicas, biodisponibilidad y metabolismo del zinc, los cuales serán abordados en la primera parte del presente artículo.
Funciones del zinc
Entre las funciones que desempeña el zinc en el organismo del cerdo podemos destacar las siguientes:
1.- Formar parte de enzimas (metaloenzimas) tales como: alcohol deshidrogenasa, fosfatasa alcalina, anhidrasa carbónica, carboxipeptidasas A y B, colagenasa, leucina aminopeptidasa, manosidasa y superóxido dismutasa.
2.- Intervenir en el metabolismo de la vitamina A, al formar parte de las enzimas retineno reductasa y alcohol deshidrogenasa.
3.- Favorecer la estabilidad de la membrana de los eritrocitos (Bettger, 1989).
4.- Jugar un papel activo en la digestión, glicolisis, síntesis de ADN, metabolismo de ácidos nucleicos y proteínas (Chesters, 1992). De ahí su importancia en la espermatogénesis como formadora de nuevas células.
5.- Participar en el metabolismo de los ácidos grasos (Cunnane y Yang, 1997). Su carencia se asocia a una disminución de ácido araquidónico circulante y a modificaciones a nivel lipídico (Samman y Roberts, 1988).
6.- Intervenir en la regulación del apetito, aunque sus mecanismos de actuación no son muy bien conocidos. En cualquier caso sí se ha podido observar que los lechones con un déficit de zinc presentaban un cuadro de anorexia, provocando una menor ganancia diaria de peso y un menor índice de transformación (Miller et al., 1968).
7.- En el verraco destaca su papel en la producción y calidad espermática, mejorando la fertilidad (Chesters y Will, 1981). Interviene en el proceso de maduración de las células de Leydig, en la respuesta a la hormona luteinizante y en la formación de esteroides (Hesketh, 1982). Además, parece influir en la estabilidad de la membrana espermática y guarda relación con el contenido proteico del plasma seminal y con el consumo de oxígeno por parte del espermatozoide.
8.- Mejorar el sistema inmunitario del cerdo ante situaciones de estrés crónico, como consecuencia de situaciones medioambientales desfavorables, debido a su participación en la maduración de los linfocitos T.
9.- Finalmente, intervenir en otras acciones relacionadas con el equilibrio ácido-base, el metabolismo de los andrógenos, a nivel de la 5a-reductasa y en la detoxicación de radicales libre.
2.- Intervenir en el metabolismo de la vitamina A, al formar parte de las enzimas retineno reductasa y alcohol deshidrogenasa.
3.- Favorecer la estabilidad de la membrana de los eritrocitos (Bettger, 1989).
4.- Jugar un papel activo en la digestión, glicolisis, síntesis de ADN, metabolismo de ácidos nucleicos y proteínas (Chesters, 1992). De ahí su importancia en la espermatogénesis como formadora de nuevas células.
5.- Participar en el metabolismo de los ácidos grasos (Cunnane y Yang, 1997). Su carencia se asocia a una disminución de ácido araquidónico circulante y a modificaciones a nivel lipídico (Samman y Roberts, 1988).
6.- Intervenir en la regulación del apetito, aunque sus mecanismos de actuación no son muy bien conocidos. En cualquier caso sí se ha podido observar que los lechones con un déficit de zinc presentaban un cuadro de anorexia, provocando una menor ganancia diaria de peso y un menor índice de transformación (Miller et al., 1968).
7.- En el verraco destaca su papel en la producción y calidad espermática, mejorando la fertilidad (Chesters y Will, 1981). Interviene en el proceso de maduración de las células de Leydig, en la respuesta a la hormona luteinizante y en la formación de esteroides (Hesketh, 1982). Además, parece influir en la estabilidad de la membrana espermática y guarda relación con el contenido proteico del plasma seminal y con el consumo de oxígeno por parte del espermatozoide.
8.- Mejorar el sistema inmunitario del cerdo ante situaciones de estrés crónico, como consecuencia de situaciones medioambientales desfavorables, debido a su participación en la maduración de los linfocitos T.
9.- Finalmente, intervenir en otras acciones relacionadas con el equilibrio ácido-base, el metabolismo de los andrógenos, a nivel de la 5a-reductasa y en la detoxicación de radicales libre.
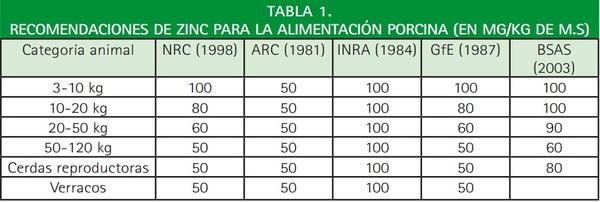
NRC: National Research Council.
ARC: Agricultural Research Council.
INRA: Institut National de la Recherche Agronomique.
GfE: Gesellschaft für Ernährungsphysiologie.
BSAS: British Society of Animal Science.
ARC: Agricultural Research Council.
INRA: Institut National de la Recherche Agronomique.
GfE: Gesellschaft für Ernährungsphysiologie.
BSAS: British Society of Animal Science.
Biodisponibilidad
Sería más correcto hablar de biodisponibilidad de la ración en su conjunto, en vez de cada una de las materias primas separadamente, aunque alguno de ellas aporte la mayor parte del zinc, por tres razones fundamentales (Windisch, 2002):
- El zinc se absorbe según las necesidades del animal.
- En el intestino el fitato forma junto con el zinc complejos no absorbibles.
- El grado de interacción del fitato con el zinc depende del nivel de cálcio de la ración.
No obstante, podemos aportar algunos datos sobre biodisponibilidad, así el zinc presente en los cereales tiene una biodisponibilidad baja para los cerdos, alrededor del 60% (Baker y Ammerman, 1995), siendo más alta en los concentrados proteicos vegetales (75-85%) (Franz et al., 1980); encontrándose la mayor biodisponibilidad en las fuentes proteicas de origen animal libre de fitatos. Si bien, en este último caso la biodisponibilidad puede disminuir cuando se mezclan con sustancias ricas en fitatos y suplementos de cálcio. En este sentido, Tucker y Salmón (1955) descubrieron que cuando los piensos a base de maíz y soja eran suplementados con un 1,5% de carbonato cálcico o fosfato cálcico o con un 2% de harina de hueso, disminuía el crecimiento y aumentaban las lesiones de paraqueratosis en cerdos. Sin embargo, todo ello se corregía cuando se suplementaba con zinc.
Por otra parte, los efectos negativos de los fitatos podrían reducirse disminuyendo el aporte del cálcio y fósforo en las raciones, aumentando la exposición a las fitasas vegetales, aportando fitasas microbianas o, bien, utilizando vitamina D3 hidroxilada para estimular la eficacia de las fitasas (Lei et al., 1993; Adeola et al., 1995 y Welch, 1997). En este sentido, debemos señalar los recientes estudios llevados a cabo por Windisch et al., (2003) quienes comprobaron como la adición de fitasas al pienso de lechones mejora la biodisponibilidad del zinc.
Metabolismo
EL zinc disponible para el cerdo depende del contenido del mismo en la ración y de la composición de la misma, pero también del porcentaje absorbido por el aparato digestivo y de la cantidad excretada.
El zinc se absorbe en función de las necesidades del organismo mediante un sistema de transporte activo, siendo la zona del yeyuno, la de mayor absorción y en menor medida la del duodeno e intestino grueso.
Cuando se añade cobre a las raciones de los cerdos, como estimulante del crecimiento, éste puede alterar la absorción del zinc. Así, cerdos en crecimiento que recibieron 250 mg de Cu/ kg de materia seca (MS), en forma de sulfato de cobre, como estimulante del crecimiento, mostraban lesiones cutáneas y un índice de mortalidad similares a las observadas en las paraqueratosis, las cuales podían ser curadas con aportes de zinc (O´Hare et al., 1960 y Suttle y Mills, 1966).

Contrariamente, la lactosa, la glucosa y los ácidos grasos insaturados aumentan la absorción del zinc.
Respecto al transporte, el zinc es transportado por el torrente sanguíneo unido mayoritariamente a la albúmina plasmática (65%). Otra parte del zinc se transporta en forma de a2-macroglobulina y en forma de restos de metalotioneína.
La mayor parte del zinc que circula por las vías sanguíneas (80%) se encuentra localizado en los hematíes (1 mg de Zn/106 hematíes), del cual el 85% se encuentra en forma de anhidrasa carbónica y un 5% en forma de cobre-zinc superóxido dismutasa.
Por otra parte, la capacidad para almacenar zinc en los tejidos, por parte de los cerdos, es escasa. En situaciones de déficit se puede movilizar zinc acumulado en los huesos, páncreas e hígado, que son los mayores depósitos de este oligoelemento (Tabla 2).
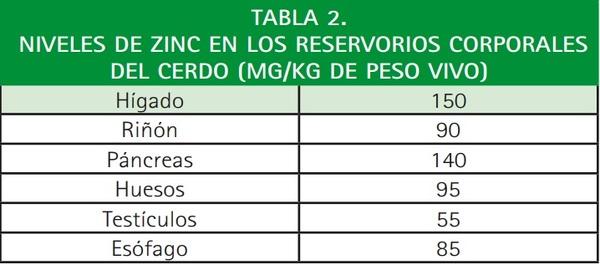
Finalmente, el zinc se elimina fundamentalmente a través de las secreciones pancreáticas y las heces, y en mucha menor medida por la orina, esperma o descamación de la piel. Se ha observado que el aporte de aminoácidos en la dieta como histidina, cistina, metionina, triptófano y lisina aumentan la eliminación urinaria del zinc. En el caso del cerdo la reabsorción está influida por la presencia de fitatos.
Necesidades de Zinc Y SUS EFECTOS SOBRE LOS LECHONES
Wedekind et al., (1994) determinaron que aproximadamente 27 mg de Zn/kg de MS eran suficientes para el crecimiento de los lechones, siempre y cuando la ración tuviera solamente 5,9 g de Ca/kg de MS y 3,6 g de fitatos/kg de MS. Cantidades similares fueron aportadas por Hill et al., (1986) para las raciones de crecimiento en cerdos (24-33 mg de Zn/kg de MS con una cantidad de cálcio de 6.5 g/kg de MS).
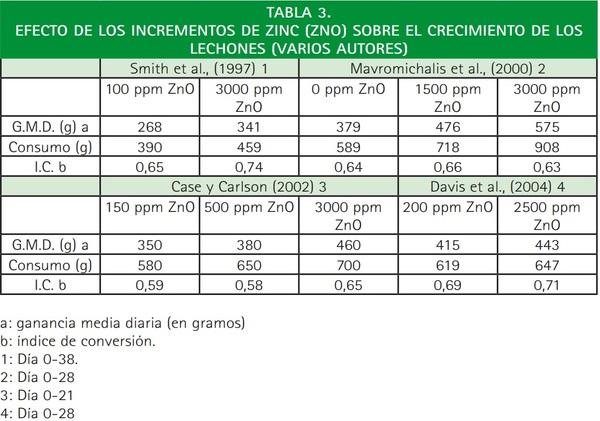
La mayoría de los autores consultados ponen de manifiesto los efectos beneficiosos de la suplementación de zinc en la ración (1500-3000 ppm) sobre el crecimiento de los lechones (Fryer et al., 1992; Tokach et al., 1992; Hahn y Baker, 1993; Schell y Kornegey, 1996; Smith et al., 1997; Mavromichalis et al., 2000; Case y Carlson, 2002; Davis et al., 2004).
La carencia prolongada del zinc en los lechones puede ser debido a aportes insuficientes en la ración, aumento de las necesidades en momentos puntuales, escasa absorción o aumento de las pérdidas por excreción, dando lugar a una serie de alteraciones bioquímicas y clínicas, entre las que podemos destacar las siguientes:
1 Estados deficitarios de zinc repercuten rápidamente en su concentración plasmática en los lechones. Si el estado deficitario persiste y se hace crónico repercute en el descenso de zinc en otros tejidos y órganos, sobre todo a nivel de páncreas e hígado (Swinkels et al., 1996).
2 Lógicamente al formar parte de varias metaloenzimas, la carencia de zinc repercute en la actividad de las mismas. En este sentido, se ha observado que ante estados carenciales de zinc en lechones disminuye la fosfatasa alcalina pero no la alcohol deshidrogenasa en el hígado (Miller et al., 1968).
3 La pérdida de apetito o anorexia es una de las manifestaciones clínicas más típicas de la deficiencia de zinc, repercutiendo en un retraso del crecimiento de los lechones (Windisch et al., 2003).
4. Cuando el déficit es muy prolongado afecta a la piel y a sus tegumentos, siendo la lesión más característica la paraqueratosis (engrosamiento, endurecimiento y agrietamiento de la piel), observándose, principalmente, en cerdos jóvenes a nivel de las extremidades y en verracos viejos. En ocasiones se pueden apreciar lesiones epiteliales en la lengua y en el esófago, similares a los estados de avitaminosis A. Estas lesiones y pequeñas heridas tardan mucho en cicatrizar, aunque ello puede ser una consecuencia indirecta del estado de anorexia.
5 Finalmente, en los lechones se aprecia una reducción en el tamaño y en la dureza del fémur, aunque ello, igualmente, puede ser debido a una disminución en la ingesta de pienso (Miller et al., 1968).
Necesidades de Zinc Y SUS EFECTOS SOBRE LOS VERRACOS
En verracos aportes de 40 mg de Zn/kg de MS pueden ser suficiente para lograr una buena producción espermática; sin embargo, ante situaciones de estrés o cuando se incrementa el número de saltos por semana, las necesidades pueden aumentar hasta 65 mg de Zn/kg de MS. E incluso, ante situaciones persistentes de estrés térmico, junto a diarreas prolongadas, que ocasionan una pérdida importante de zinc en heces, las necesidades de zinc pueden llegar a 100 mg de Zn/kg de MS (80-120 mg de Zn/kg de MS) (Close y Roberts, 1993).
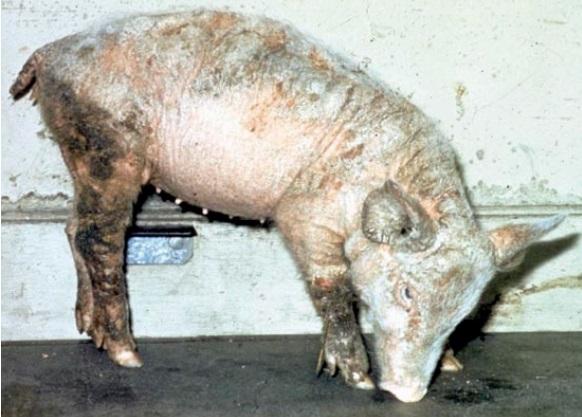
Respecto, a las consecuencias patológicas que origina su deficiencia diremos que a nivel de los verracos se detecta una pérdida del apetito y, consecuentemente, una reducción del peso, falta de líbido, impotencia e hipofertilidad. Se reduce el peso de los testículos, disminuyendo la concentración de zinc en los mismos, próstata y plasma seminal, con degeneración histológica del epitelio espermático a nivel de los túbulos seminíferos y células de Leydig (Bunce, 1989). Así mismo, puede ocasionar un retraso en la pubertad, con atrofia del timo, órganos linfoides y bazo. Por otra parte, se observa una disminución del sistema inmunológico (Chesters y Will, 1981).

Todas estas alteraciones bioquímicas y evidencias clínicas ayudan al veterinario al correcto diagnóstico del déficit de zinc. En cualquier caso, la concentración de zinc en suero o plasma es el indicador más utilizado en el diagnóstico aunque su rápida respuesta suponga ciertos límites de precisión y sensibilidad como criterio de diagnóstico. Los valores normales de zinc pueden oscilar entre 0,8 y 1,2 mg de Zn/l (12,3- 18,5 µmol).
También se puede utilizar como criterio de diagnóstico la reducción en el plasma de la fosfatasa alcalina aunque hemos de tener en cuenta que también puede estar disminuida por alteraciones óseas e intestinales (Swinkels et al., 1996).
Suplementación Y TOXICIDAD DEL ZINC
A la hora de suplementar la ración de los cerdos con zinc podemos utilizar fuentes inorgánicas u orgánicas. Entre las primeras las más utilizadas se encuentran el óxido de zinc y el sulfato de zinc de calidad alimentaria. Mientras que entre las fuentes orgánicas destacamos los complejos Zn:metionina, Zn:lisina y Zn:picolinato, los cuales actúan protegiendo al zinc frente a los antagonistas naturales como son los fitatos.
No está totalmente claro cual de ambas fuentes es mejor a la hora de suplementar el pienso al ganado porcino, ya que los estudios de Hill et al., (1986) no detectaron ninguna diferencia entre la administración de Zn:metionina (con o sin la incorporación de ácido picolínico) y la administración de sulfato de zinc, cuando eran añadidos a raciones basales en cerdos en crecimiento. Igualmente, Swinkels et al., (1996) no encontraron ninguna diferencia entre fuentes inorgánicas (sulfato de zinc) y fuentes orgánicas (Zn:aminoácidos), ambas aportaban 45 mg de Zn a un pienso que contenía 17 mg de Zn/kg de MS.

Sin embargo, estudios llevados a cabo por Wedekind et al., (1994) y Schell y Kornegay (1996) sobre raciones a base de soja y maíz en lechones, determinaron que la biodisponibilidad del zinc variaba en función de la fuente, estableciendo el siguiente orden de mayor a menor biodisponibilidad: sulfato de zinc > Zn:metionina > Zn:lisina > óxido de zinc. Recientemente, Windisch (2005) ha demostrado que cuando se utilizan dietas que contienen fitatos, el potencial de absorción de zinc como sulfato es menor que el del zinc derivado de compuestos orgánicos.
Finalmente, destacamos que las últimas investigaciones ponen de manifiesto una mayor tasa de crecimiento post-destete con fuentes orgánicas que con óxido de zinc (Poulsen y Carlson, 2001; Carlson y Poulsen, 2003).
En cuanto a la toxicidad del zinc, diremos que en líneas generales el cerdo suele ser tolerante a consumos excesivos, aunque ello depende del contenido de cálcio,
cobre, hierro y cadmio de la ración. Brink et al., (1959) determinaron que ingestas de 1 g de Zn/kg de MS en forma de carbonato no producían ninguna alteración en lechones en transición; ahora bien, cuando se incrementaba el aporte hasta 4-8 g de Zn/kg de MS apreciaban un incremento de la mortalidad. Sin embargo, estas últimas cantidades (4 g de Zn/kg de MS) no resultan tan perjudiciales si el contenido en cálcio de la ración se mantiene entre 7 y 11 g de Ca/kg de MS (Hsu et al., 1975).
Por su parte, Smith et al., (1997) detectaron que, incluso, cantidades ligeramente inferiores a 3 g de Zn/kg de MS en forma de óxido de zinc, pueden estimular el crecimiento de los lechones sin efectos tóxicos; es más, Zhang et al., (1995) llegaron a proponer el consumo elevado de zinc para controlar la disentería porcina.
RECOMENDACIONES PRÁCTICAS
El riesgo de deficiencias de zinc en cerdos disminuye a medida que aumentamos la proporción de proteínas de origen animal, tales como la harina de carne y pescado y subproductos lácteos (caseína) en el pienso. Igualmente, también es mínimo el riesgo siempre que no sean anormalmente altos los contenidos de cálcio y cobre de la ración, de ahí la importancia del equilibrio mineral de la ración, o bien, los contenidos en fitatos.
En cualquier caso las deficiencias de zinc pueden prevenirse con facilidad en cerdos mediante la simple incorporación de sales minerales (preferiblemente en forma de sulfato de zinc) o fuentes orgánicas de zinc al corrector mineral normalmente utilizado. Aportes de 50 mg de Zn/kg de MS son suficiente, excepto para aquellos cerdos que reciban sulfato de cobre como estimulante del crecimiento, en estas circunstancias debemos aportar hasta tres veces más.
Otro aspecto a tener en cuenta es intentar mejorar la biodisponibilidad del zinc, aumentando la actividad de las fitasas, para contrarrestar uno de sus principales enemigos como son los fitatos.
Finalmente, recomendamos en situaciones de estrés térmico (> 32º C), para mejorar la fertilidad de los verracos, aumentar el aporte de zinc hasta 100 mg de Zn/kg de MS, junto con un incremento de vitamina C.
Articulo extraido de la Biblioteca Digital del Ministerio de Agricultura, Pesca y Alimentación del Gobierno de España: Efecto del zinc en la alimentación porcina, publicado en engormix.com por gentileza del autor
Referencias
- Adeola, O.; Lawrence, B.V.; Sutton, A.C. y Cline, T.R. 1995. Phytase-induced changes in mineral utilisation in zinc-supplemented diets for pigs. Journal of Animal Science, 73: 3384-3391.
- Baker, D.H. y Ammerman, C.B. 1995. Zinc bioavailability. En: Ammerman, C.B.; Baker, D.H y Lewis, A.J. (Eds) Bioavailability of nutrients and carbonic anhydrase in lambs. Indian Journal of Animal Science, 43: 140-144.
- Bettger, W.J. 1989. The effect of dietary zinc deficiency on erythrocyte-free and membrane-bound amino acids. Nutrition Research, 9:911-919.
- Brink, M.F.; Becker, D.E.; Terrill, S.W. y Jensen, A.H. 1959. Zinc toxicity in the wealing pig. Journal of Animal Science, 18: 836-842.
- Bunce, G.E. 1989. Zinc in endocrine function. Mills C.F. Ed. Springer Verlag, Londres, 249-258.
- Carlson, D. y Poulsen, H.D. 2003. Dietary zinc enhances gastrointestinal function in pigs after weaning. Proceedings of the 9th
- International Symposium on Digestive Physiology of Pig, Banff, AB, Canada, May 14-18, Vol. 2, pp. 227-229.
- Case, C.L. y Carlson, M.S. 2002. Effect of feeding organic and inorganic sources of additional zinc on growth performance and zinc balance in nursery pigs. J. Anim Sci., 80: 1917-1924.
- Chesters, J.K. 1992. Trace element-gene interactions. Nutrition Reviews, 50: 217-223.
- Chesters, J.K. y Will, M. 1981. Measurement of zinc flux through plasma in normal and endotoxin stressed pigs and the effects of Zn supplementation during stress. British Journal of Nutrition, 46: 119-130.
- Close, W.H. y Roberts, F.G. 1993. Nutrition of the working boar. Recent Developments in pig nutrition 2. Ed. D.J. A. Cole; W. Haresing y P.C. Garnsworhty. Nottingham University, Press, pp: 347-368.
- Cunnane, S.C. y Yang, J. 1997. Disruption of the metabolism of polyunsatured fatty acids (PUFA) during moderate zinc deficiency. En: Fischer, P.W.F.; L´Abbe, M.R.; Cockell, K.A. y Gibson, R.S. (Eds). Trace element in man and animals. 9 Proceedings of the ninth International Symposium. NRC. Research Press, Ottawa, Canada, pp: 604-608.
- Davis, M.E.; Brown, D.C.; Maxwell, C.V.; Johnson, Z.B.; Kegley, E.B. y Dvorak, R.A. 2004. Effect of phosphorylated mannans and pharmacological additions of zinc oxide on growth and immunocompetence of weanling pigs. J. Anim Sci., 82:581-587.
- Franz, K.B.; Kennedy, B.M. y Fellers, D.A. 1980. Relative bioavailability of zinc from selected cereals and legumes using rat growth. Journal of Nutrition, 110: 2272-2283.
- Fryer, A.; Miller, E.R.; Ku, K. y Ullrey, D.E. 1992. Effect of elevated dietary zinc on growth performance of weanling swine. Michigan State Univ. Rep of Swine Res., 520: 128-133.
- Han, J.D. y Baker, D.H. 1993. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci., 71: 3020-3024.
- Hesketh, J.E. 1982. Effects of dietary zinc deficiency on Leydig cell ultrastructure in the boar. Journal of Comparative Pathology, 92:239-247.
- Hill, D.A.; Peo, E.R.; Ledwis, A.J. y Crenshaw, J.D. 1986. Zinc-amino acid complexes for swine. Journal of Animal Science, 63: 121-130.
- Hsu, F.S.; Krook, L.; Pond, W.G. y Duncan, J.R. 1975. Interactions of dietary calcium with toxic levels of lead and zinc in pigs. Journal of Nutrition, 105: 112-118.
- Lei, X.G.; Ku, P.K.; Miller, R.E.; Ullrey, D.E. y Yokoyama, M.T. 1993. Supplental microbial phytase improved bioavailability of dietary zinc to weanling pigs. Journal of Nutrition, 123: 1117-1123.
- Mavromichalis, I.; Peter, C.M.; Parr, T.M.; Ganessunker, D. y Baker, D.H. 2000. Growth-promoting efficacy in young pigs of two sources of zinc oxide having either a high or a low bioavailability of zinc. J. Anim Sci., 78: 2896-2902.
- Miller, E.R.; Luecke, R.W.; Ullrey, D.E.; Baltzer, B.V.; Bradley, B.L. y Hoefer, J.A. 1968. Biochemical skeletal and allometric changes due to zinc deficiency in the baby pig. Journal of Nutrition, 95: 278-286.
- O´Hare, P.J.; Newman, A.P. y Jackson, R. 1960. Parakeratosis and copper poisoning in pigs fed a copper suplement. Australian Veterinary Journal, 36: 225-229.
- Poulsen, H.D. y Carlson, D., 2001. Bioavailability of zinc from different zinc sources. 52nd Annual Meeting of the European Association for Animal Production, Budapest, Hungary. Book of Abstracts EAAP, Wageningen Pers, p. 123.
- Samman, S. y Roberts, D.C.K. 1988. Zinc and cholesterol metabolism. Nutrition Research, 8: 559-570.
- Schell, T.C. y Kornegay, E.T. 1996. Zinc concentration in tissues and performance of weanling pigs fed pharmacological levels of zinc from ZnO, Zn-Methionine, Zn-Lysine or ZnSO4. J. Anim. Sci., 74: 1584-1593.
- Smith, J.W.; Tokach, M.D.; Goddband, R.D.; Nelssen, J.L. y Richert, B.T. 1997. Effects of the interrelationship between zinc oxide and copper sulfate on growth performance of early-weaned pigs. J. Anim. Sci., 75: 1861-1866.
- Suttle, N.F. 1983. Assessment of the mineral and trace element status of feeds. En: Roberds, G.E. y Packham, R.G. (eds). Proceedings of the
- Second Symposium of the International Network of Feed Information Centres. Commonwealth Agriculture Bureaux, Farnham Royal, UK, pp: 211-237.
- Suttle, N.F. y Mills, C.F. 1966. Studies on the toxicity of copper to pigs. 1. Effects of oral suplements of zinc and iron salts on the development of copper toxicosis. British Journal of Nutrition, 20: 135-148.
- Swinkels, J.W.; Kornegay, E.T.; Zhou, W.; Lindemann, M.D.; Webb, K.E. y Verstegen, M.W.A. 1996. Effectiveness of a zinc amino acid chelate and zinc sulphate in restoring serum and soft tissue concentrations when fed to zinc-depleted pigs. Journal of Animal Science, 74: 2420-2430.
- Tokach, L.M.; Tokach, M.D.; Goddband, R.D.; Nelssen, J.L.; Henry, S.C. y Marsteller, T.A. 1992. Influence of zinc oxide in starter diets on pig performance. American Association of Swine Practitioners Proceedings, p. 411.
- Tucker, H.F. y Salmon, W.D. 1955. Parakeratosis of zinc deficiency disease in the pigs. Proceedings of the Society for Experimental Biology and Medicine, 88: 613-616.
- Wedekind, K.J.; Lewis, A.J.; Giesemann, M.A. y Miller, P.S. 1994. Bioavailability of zinc from inorganic and organic sources for pigs fed corn-soybean meal diets. Journal of Animal Science, 72: 2681-2689.
- Welch, R.M. 1997. Trace elementinteractions in food crops. En: Fischer, P.W.F.; L´Abbe, M.R.; Cockell, K.A. y Gibson, R.S. (Eds). Trace elementin man and animals. 9 Proceedings of the ninth International Symposium. NRC. Research Press, Ottawa, Canada, pp: 6-9.
- Windisch, W. 2002. Interaction of chemical species with biological regulation of the metabolism of essential trace elements. Anal. Bioanal. Chem., 372: 421-425.
- Windisch, W. 2005. Biovailibility of Zn from sulfat, glycine complex and proteinate in 65Zn labelled rats fed phytate-enriched diets. J. Anim. Physiol. Anim. Nutr. (en prensa).
- Windisch, W.; Broz, J y Roth, F.X. 2003. Effect ofmicrobial phytase onthe biovailibility of zinc inpiglet diets. En: Proceedings of the Society of Nutrition Physiology. Breves, G. Ed. Frankfurt/Main, Germany: DGL-Verlags GmbH, p. 33.
- Zhang, P.; Duhamel, G.E.; Mysore, J.V. Carlson, M.P. y Schneider, N.R. 1995. Prophylactic effect of dietary zinc ina laboratory mouse model of swine dysentry. Journal of Veterinaty Research, 56: 334-339
Temas relacionados
Autores:

Referentes que Recomendaron :
Jose Ángel Ayala Cutillas, Marco Antonio Jacho LópezÚnete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicación
Universidad de Guadalajara
6 de marzo de 2022
Excelente gracias por su aporte




Te puede interesar
.jpg&w=3840&q=75)
Provimi Argentina