
Contenido patrocinado por:
Biotecno
Bases biológicas de las enzimas
Publicado: 18 de marzo de 2021
Por: Nathalia María del Pilar Correa Valencia, MV, MSc, DSc.
Enzimas: Bases biológicas
Desde el punto de vista investigativo, se ha logrado un desarrollo relativamente rápido en la determinación de las secuencias y estructuras de las proteínas. Esto, en combinación con el conocimiento de miles de funciones enzimáticas y cientos de mecanismos químicos y bioquímicos, hace posible que hoy en día la combinación en amplitud y profundidad de nuestra comprensión acerca de la evolución de las enzimas. En este mismo sentido, la predicción de la función proteica ha generado una variedad de métodos prometedores que ofrecen la posibilidad de identificar enzimas en toda gama de funciones químicas y sobre numerosas especies. Este conocimiento es esencial en la comprensión de las bases de la resistencia a los antibióticos (e.g. betalactamasas), así como en la ingeniería de proteínas y en el diseño de enzimas de novo.
Se presenta entonces una no tan nueva disciplina, la enzimología, que aunque se deriva fundamentalmente a partir de la bioquímica, involucra otras ciencias como la filogenética, genómica, química orgánica, química computacional, termodinámica y metabolómica.
¿Qué son las enzimas y cuál es su estructura?
“Todas las enzimas son proteínas pero no todas las proteínas son enzimas”, ha sido una premisa de la biología; sin embargo, actualmente se considera que las enzimas son comúnmente de naturaleza proteica, pero también existen enzimas como las ribozimas que debaten dicha condición.
Las enzimas son moléculas orgánicas que actúan como catalizadores de reacciones químicas, es decir, aceleran la velocidad de reacción, disminuyendo a su vez la energía de activación (ΔG). Al modificar la misma, logran procesos energéticamente favorables, actuando sobre los sustratos de reacción y facilitando su conversión en productos necesarios en los sistemas orgánicos. Adicionalmente, es importante mencionar que las enzimas son extremadamente selectivas con sus sustratos y reacciones. Esto es tan definitivo que el conjunto de enzimas de una célula —que viene determinado por lo genes que expresa, fija el tipo de metabolismo celular predominante.
Casi todos los procesos en las células requieren de enzimas, logrando tasas significativamente más rápidas. Las enzimas no alteran el balance energético, ni modifican el equilibrio de la reacción. Es así como una reacción catalizada por una enzima alcanza el mismo equilibrio, pero mucho más rápido, que la misma reacción no catalizada.
Cuando los bioquímicos se dieron cuenta de que las enzimas tenían una estructura persistente y que la destrucción de esa estructura podría abolir la actividad enzimática, rápidamente se adoptó la opinión de que las enzimas eran andamios rígidos cuya especificidad y poder catalítico se derivaban de la forma de ajuste del sustrato adecuado en la superficie preformada e inflexible de la enzima, así como una llave encaja en una cerradura. Hace 50 años, Daniel Koshland desafió este punto de vista, proponiendo que la superficie de la enzima era flexible y que solo el sustrato específico induciría la(s) adecuada(s) interacciones que llevarían a la catálisis.
Si miramos con atención la estructura tridimensional de las enzimas que regularmente se muestra en las publicaciones científicas, es difícil imaginar que aún hay gente viva a la fecha que recuerda cuando muchos bioquímicos pensaron que las enzimas no tenían una estructura ordenada. Este fue el caso hasta que James Sumner cristalizó la ureasa (enzima con la capacidad de descomponer la urea en carbonato de amonio) en 1926 —un desarrollo tan revolucionario que él fue puesto bajo custodia porque se consideraba que era un lunático peligroso cuando trató de explicar lo que le había hecho.
Las implicaciones del descubrimiento de Sumner de la ureasa fueron profundas; hasta entonces, no se sabía que las enzimas eran proteínas en su gran mayoría. Veinte años después, Sumner ganó el Premio Nobel de Química de 1946 "por su descubrimiento de que las enzimas pueden cristalizarse", un descubrimiento que sentó las bases para la primera estructura cristalina de rayos X de una enzima, la lisozima de clara de huevo de gallina, en 1965. A hoy tenemos certezas en cuanto a la estructura flexible, adaptable y tridimensional de las enzimas.
¿Cómo se denominan y clasifican las enzimas?
En forma sistemática, el nombre de una enzimase construye con la siguiente información: Nombre del sustrato (o sustratos), nombre del cambio químico que realiza la enzima sobre el sustrato y el sufijo asa.
Por ejemplo, tenemos la siguiente reacción mediada por enzima:

Nombre de los sustratos: NADH (dinucleótido de nicotinamida y adenina o nicotinamida adenina dinucleótido, coenzima que participa en el Ciclo de Krebs) y Coenzima Q (antioxidante endógeno).
Cambios químicos que realiza la enzima: Oxida al NADH y reduce a la Coenzima Q
Por lo que el nombre sistemático de esta enzima es NADH, Coenzima Q óxido-reductasa.
En 1964 la Comisión de Enzimas (EC) de la Unión Internacional de Bioquímica y Biología Molecular (IUBMB; https://iubmb.org/) propuso una nomenclatura, clasificándolas en seis clases, de tal forma que cada enzima debería quedar identificada con un nombre completo no ambiguo y con un código numérico de cuatro partes, separadas por puntos. Para la primera parte tenemos la siguiente clasificación:
Clase 1: Óxido-reductasas (e.g. deshidrogenasas, oxidasas, oxigenasas, peroxidasas, reductasas), encargadas de catalizar las reacciones celulares donde se pierden y se ganan electrones.
Clase 2: Transferasas (e.g. fosfotransferasas, aminotransferasas, metiltransferasas, glicosiltransferasas), que catalizan aquellas reacciones celulares donde un grupo de átomos se transfiere de un sustrato a otro.
Clase 3: Hidrolasas (e.g. lipasas-esterasas, glicosidasas, proteinasas, fosfatasas), catalizan la ruptura de un gran número de biomoléculas usando como co-sustrato a la molécula de H2O. Incluyen las lipasas sublinguales, gástricas y pancreáticas, α- y β-amilasas y las proteinasas.
Clase 4: Liasas (e.g. descarboxilasas-carboxiliasas, deshidratasas-hidroliasas, desaminasas-amonioliasas), que generan rupturas no oxidativas, no reductivas y no hidrolíticas en las moléculas.
Clase 5: Isomerasas (e.g. racemasas, epimerasas, mutasas, cis-trans isomerasas, isomerasas de grupo funcional), que catalizan conversiones entre los diferentes tipos de isómeros, estructurales, geométricos y ópticos.
Clase 6: Ligasas (e.g. carboxilasas, sintetasas). Se agrupan aquí las enzimas que catalizan la formación de nuevas sustancias, ya sea agregando a un sustrato grupos como CO2, NH3, HSCoA, o condensando varias sustancias para generar otra diferente. Tales síntesis y condensaciones requieren el consumo de energía, expresamente contenida en un nucleótido como ATP y GTP.
Clase 2: Transferasas (e.g. fosfotransferasas, aminotransferasas, metiltransferasas, glicosiltransferasas), que catalizan aquellas reacciones celulares donde un grupo de átomos se transfiere de un sustrato a otro.
Clase 3: Hidrolasas (e.g. lipasas-esterasas, glicosidasas, proteinasas, fosfatasas), catalizan la ruptura de un gran número de biomoléculas usando como co-sustrato a la molécula de H2O. Incluyen las lipasas sublinguales, gástricas y pancreáticas, α- y β-amilasas y las proteinasas.
Clase 4: Liasas (e.g. descarboxilasas-carboxiliasas, deshidratasas-hidroliasas, desaminasas-amonioliasas), que generan rupturas no oxidativas, no reductivas y no hidrolíticas en las moléculas.
Clase 5: Isomerasas (e.g. racemasas, epimerasas, mutasas, cis-trans isomerasas, isomerasas de grupo funcional), que catalizan conversiones entre los diferentes tipos de isómeros, estructurales, geométricos y ópticos.
Clase 6: Ligasas (e.g. carboxilasas, sintetasas). Se agrupan aquí las enzimas que catalizan la formación de nuevas sustancias, ya sea agregando a un sustrato grupos como CO2, NH3, HSCoA, o condensando varias sustancias para generar otra diferente. Tales síntesis y condensaciones requieren el consumo de energía, expresamente contenida en un nucleótido como ATP y GTP.
El código numérico asignado a las enzimas esuna identificación derivada de la clasificación, que contiene cuatro partes separadas por puntos, así: X. Y. Z. W. El número en la posición X denota la clase a la cual pertenece la enzima (indica el cambio químico global que realiza la enzima sobre el sustrato). El número en la posición Y denota la subclase (indicando un cambio más específico en la transformación del sustrato, como por ejemplo, el grupo funcional que se oxida, reduce, transfiere, trasforma, o el enlace que se destruye). El número en la posición Z denota a subsubclase (indicando una aún mayor especificidad de los cambios químicos señalados por el número en Y, como por ejemplo, sobre cuáles átomos se realizan las oxidaciones o reducciones, o cuales grupos de átomos se desprenden o qué enlaces se forman). Y finalmente, el número en la posición W corresponde a la identificación precisa de las sustancias sobre las cuales actúa la enzima y normalmente indica el orden en el que cada enzima se va agregando a la lista.
Por ejemplo, la enzima tripsina (enzima digestiva) tiene como identificación el código EC 3.4.21.4, lo cual significa que es una hidrolasa (clase 3), que rompe con agua un enlace peptídico (subclase 4), usando para ello un residuo de serina en su centro activo (subsubclase 21). Esta enzima es la cuarta (4) agregada a una lista de proteinasas.
A 2017 se tenían identificadas 6.028 ECs. En sentido estricto, la EC clasifica las reacciones enzimáticas, no las enzimas, pero cada número de EC suele estar vinculado a todas las secuencias enzimáticas que catalizan esa reacción, incluso si los mecanismos enzimáticos difieren.
Adicionalmente, de forma histórica las proteínas se han unificado en familias basadas en similitud de secuencia de aminoácidos. Las familias de proteínas se combinaron en superfamilias basadas en actividades catalíticas similares, motivos y otras características conservadas. Se puede inferir que las proteínas dentro de la misma superfamilia pueden haber evolucionado a partir de un antepasado común, incluso pueden tener actividades enzimáticas dramáticamente diferentes o ninguna actividad (conocida) en absoluto (Galperin & Koonin, 2012)
¿Cómo se estructuran y cómo funcionan las enzimas?
Las estructuras tridimensionales de las enzimas son una fuente de información mucho más rica para comprender la función que la secuencia sola. Por el momento, se conocen aproximadamente 134.000 estructuras diferentes en las enzimas que se han secuenciado y que posteriormente se han analizado a partir de química computacional, permitiendo la identificación de mecanismos funcionales y de vínculos evolutivos distantes conservados (https://www.ebi.ac.uk/thornton-srv/m-csa/; Mechanism and Catalytic Site Atlas, M-CSA).
De forma general, se sabe que los sustratos se unen al centro activode la enzima, que es la parte dónde tiene lugar la catálisis (proceso por el cual se aumenta la velocidad de una reacción química, debido a la participación de una sustancia llamada catalizador), formada por un sitio de unióndel sustrato y un sitio catalítico. Tras la unión del sustrato al centro activo se forman los productosde la reacción.
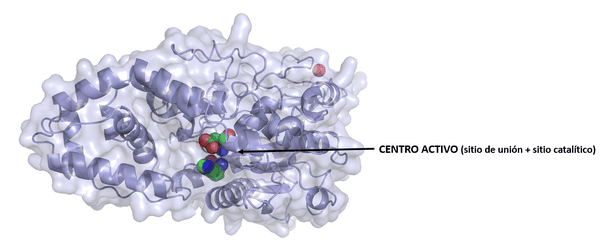
Al igual que otros catalizadores, las enzimas no se consumen en las reacciones que catalizan. Aunque existen distintos grados de especificidad, las enzimas son muy específicos, y en general más que otros catalizadores, dado que debe existir gran diversidad de enzimas para catalizar alrededor de 4.000 reacciones bioquímicas
También existen enzimas diseñadas y producidas artificialmente, que al igual que los naturales, tienen usos industriales y comerciales importantes. Por ejemplo, en la síntesis de antibióticos, de productos de limpieza, la fabricación de alimentos, la industria textil o la producción de biocombustibles.
En cuanto al proceso, la reacción enzimática requiere un aporte inicial de energía, que debe ser igual o mayor a lo que se denomina energía de activación(EA), de forma que cuanto menor es la EA más fácil es que ocurra la reacción. Los catalizadores actúan disminuyendo la EA. Las enzimas son catalizadores biológicos, más eficaces que los catalizadores inorgánicos, pudiendo disminuir la EA hasta 15 veces más.
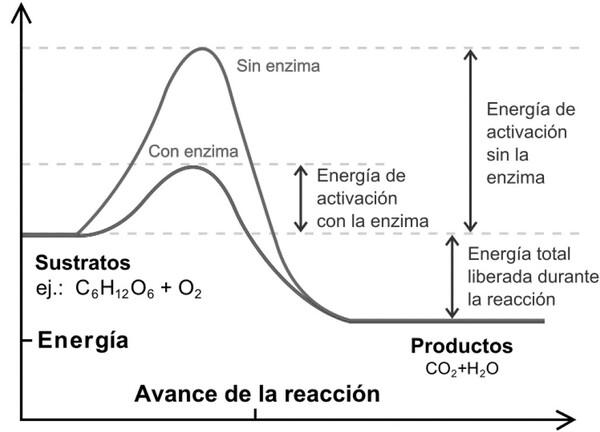
En la gráfica, la ecuación química C6H12O6 + O2 → CO2 + H2O, representa la degradación de la glucosa presente en los alimentos que se ingieren y que son degradados o desdoblados en el cuerpo para proporcionar la energía necesaria para cumplir con las funciones vitales.
Para que una reacción química tenga lugar, las moléculas de los sustratos deben chocar con una energía y una orientación adecuadas. Al unirse al centro activo de la enzima, los sustratos adquieren la orientación óptima para la reacción y se modifican sus propiedades químicas, debilitando los enlaces y facilitando la formación de otros nuevos. Estos dos eventos son el mecanismo por el cual el enzima disminuye la EA. Existen dos modelos para explicar cómo el sustrato se une al centro activo de la enzima: el modelo llave-cerradura y el modelo del ajuste inducido: El modelo de llave-cerradurasupone que la estructura del sustrato (llave) y la del centro activo del enzima (cerradura) son complementarias. Este modelo explica el mecanismo en algunos casos, pero no es siempre correcto. En algunos casos, el centro activo adopta la conformación catalítica sólo cuándo se une el sustrato (modelo del ajuste inducido). Según este modelo la unión del sustrato al centro activo provoca el cambio conformacional que da lugar a la formación del producto.
¿Qué fáctores regulan/afectan su función biológica?
La actividad de las enzimas puede ser afectada por otras moléculas, además de otros factores físico-químicos, como la temperatura, pH, cofactores y coenzimas, inhibidores (reversibles, irreversibles), y la misma geometría (orientación y proximidad al sustrato).
El aumento de la temperatura en general, aumenta a su vez la velocidad de las reacciones químicas. Las reacciones catalizadas por enzimas siguen esta norma, pero a partir de cierta temperatura se desnaturaliza la enzima. La temperatura a la cual la actividad catalítica es máxima se llama temperatura óptima. Por encima de esa temperatura la actividad enzimática disminuye con el aumento de temperatura hasta desaparecer.
Las enzimas son generalmente proteínas globulares que pueden presentar tamaños muy variables. Como proteínas que son, poseen grupos ionizables en las cadenas laterales de los aminoácidos que los forman. El estado de ionización (carga positiva, negativa o neutra) de estos grupos funcionales, y por lo tanto, la conformación estructural de la enzima, va a depender del pH del medio. Por este motivo, la conformación será más adecuada para la catálisis, y por lo tanto la actividad será óptima a un pH determinado, denominado pH óptimo. La mayoría de enzimas son muy sensibles a los cambios de pH, y pequeñas variaciones pueden provocar su desnaturalización, por lo que en los sistemas biológicos existen los llamados amortiguadores de pH o tampones.
Casi la tercera parte de las enzimas requieren de la presencia de sustancias no proteicas que participan en la acción catalítica, los cofactores. Muchos son cationes inorgánicos como el Fe2+, Mg2+, Mn2+, Zn2+, o Cu2+. Si el cofactor es una molécula orgánica se denomina coenzima. Cuando los cofactores y las coenzimas se encuentran unidos covalentemente a la enzima se llaman grupos prostéticos. La forma catalíticamente activa de la enzima, es decir, el enzima unido a su grupo prostético, se llama holoenzima, siendo su parte proteica la apoenzima). Muchas coenzimas son o se sintetizan a partir de vitaminas. Por ejemplo, la vitamina B1 o tiamina, cuyo derivado, el pirofosfato de tiamina, es esencial para el metabolismo energético de los glúcidos.
Los inhibidores enzimáticos son moléculas que disminuyen o impiden la actividad de las enzimas, mientras que los activadores son moléculas que incrementan su actividad. Muchos fármacos son moléculas inhibidoras, activadoras o antagonistas de enzimas. Los inhibidores enzimáticos actúan ocupando temporalmente el centro activo por semejanza estructural con el sustrato (inhibidor competitivo) o alterando la conformación espacial de la enzima, de forma que no pueda unirse al sustrato (inhibidor no competitivo).
Dado que son proteínas, las enzimas presentan una estructura tridimensional de la que depende su actividad biológica (estructura nativa), y esa actividad se verá afectada por cambios conformacionales. La pérdida de la estructura nativa (desnaturalización) conlleva la pérdida de la actividad catalítica, y puede ser reversible o irreversible, dependiendo del grado en que se produzca. Son agentes desnaturalizantes el calor, la presión, los valores de pH extremos, altas concentraciones de sales, detergentes y agentes caotrópicos. Sin embargo, también se producen pequeños cambios conformacionales durante la catálisis, sin llegar a perder la estructura nativa ni la actividad. Por ejemplo, el acoplamiento enzima-sustrato y la catálisis pueden implicar cambios conformacionales encaminados a que se produzca la reacción.
Las enzimas son catalizadores fisiológicos esenciales involucrados en todos los procesos de la vida, incluido el metabolismo, la señalización celular y la motilidad, así como el crecimiento y división. Son dianas farmacológicas atractivas debido a la presencia de bolsas de unión de sustrato definidas, que se pueden explotar como sitios de unión para inhibidores de enzimas farmacéuticas. Entender los mecanismos de las enzimas y el modo de acción molecular de inhibidores es indispensable para el descubrimiento y desarrollo de fármacos eficaces y seguros, potentes y novedosos. Así mismo, la combinación de conceptos clásicos con nuevos métodos experimentales y de análisis de datos definirá nuevas y necesarias rutas de investigación e innovación en este interesante tema.
Temas relacionados
Autores:

Referentes que Recomendaron :
Sebastian Puerta Atehortua, Olga Lucia Quevedo Únete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicaciónTe puede interesar
.jpg&w=3840&q=75)