Aplicaciones de los minerales traza en la nutrición de cerdos
Publicado: 9 de mayo de 2011
Por: Federico Salvador y Nora Castillo, Universidad Autónoma de Chihuahua, Chihuahua, México
Introducción.
Los minerales traza son comúnmente ignorados como fuente de nutrientes en los alimentos de los animales. Sus roles fisiológico son frecuentemente subestimados, sin embargo, ellos son necesarios para mantener funciones del cuerpo, optimizar el crecimiento, reproducción y respuesta inmune. Por lo que ellos juegan una parte importante en la determinación del estado de salud del animal. Una Deficiencia de estos elementos traza puede causar una considerable reducción en el comportamiento, y por lo general los ingredientes del alimento son deficientes de éstos oligoelementos (Close, 2000).
Requerimientos de minerales.
Es difícil justificar los requerimientos para minerales en la misma manera como lo es para energía, proteína y aminoácidos. La mayoría de los trabajos relacionados con los requerimientos minerales fueron realizados en los años 1960 y 1970 y por consiguiente puede no aplicar para el animal moderno. Esto es evidente en la reciente revisión del NRC (1998), en la cual, con pocas excepciones, muestra pocas diferencias en los requerimientos propuestos por el NRC (1988) o ARC (1981) (Tabla 1). Las diferencias en los requerimientos de nutrientes son el resultado de los objetivos de producción y del estado fisiológico del animal. Se ha sugerido que los requerimientos minerales para el crecimiento en los cerdos híbridos modernos de rápido crecimiento son cerca del doble del nivel requerido para los cerdos de lento crecimiento de unos 20 o 30 años atrás (Close, 2002).
Tabla 1. Requerimientos en la dieta para elementos traza (mg/Kg de dieta).
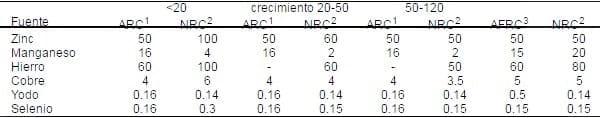 1 ARC (1981) por Kg. de material seca
1 ARC (1981) por Kg. de material seca2 NRC (1998) 90% material seca
3 AFRC (1990) 90% de material seca
La importancia de los minerales traza es grande, y aunque, están incluidos tradicionalmente en la dieta en cantidades muy pequeñas en forma de premezclas de sales inorgánicas tales como los sulfatos, cloruros carbonatos y óxidos, hay varios factores que pueden reducir su disponibilidad cuando son ingeridos por los animales. Algunos investigadores de la nutrición animal han mostrado que la disponibilidad de algunos minerales traza puede ser mejorada por la unión de ellos, con ligandos orgánicos. Por esta razón, el número de minerales traza orgánicos comerciales ha aumentado y también su uso en la industria del cerdo. En vista del incrementado uso de productos por la industria de la alimentación animal es interesante revisar brevemente las funciones bioquímicas esenciales (Close, 2002).
Química biológica de metales traza orgánicos
De los elementos traza como arsénico (As), boro (B), cromo (Cr), cobalto (Co), Cobre (Cu), Yodo (I), Flúor (F), hierro (Fe), manganeso (Mn), molibdeno (Mb), niquel (Ni), selenio (Se), Sílice (Si), vanadio (V) y zinc (Zn) (Power y Horgan, 2000), los de interés en la nutrición del cerdo por sus benéficos en forma orgánica son Cr, Se, y Cu.
Se han definido varias categorías de los minerales traza orgánicos usados en la práctica en la agricultura por la Association of American Feed Control Officials (AAFCO, 1998). Éstas son mostradas en la Tabla 2.
Tabla 2. Definiciones AAFCO para los complejos minerales orgánicos.
| Complejo metal aminoácido | Es el producto resultante del complejo de una sal metálica soluble con un aminoácido (s). |
| Quelato metal amino | Es el producto resultante de la reacción de un ión metálico de una sal metálica soluble con aminoácidos en la siguiente proporción, un mol metálico a uno o tres (preferiblemente dos) moles de aminoácidos para formar uniones covalentes. El peso promedio de los aminoácidos hidrolizados pueden ser de aproximadamente 150 y el peso molecular resultante de un quelato no excede mas de 800 |
| Complejo metálico polisacárido | Es el producto resultante de la unión de una sal soluble con un polisacárido |
| Proteinato metálico | Es el producto resultante de la quelación de una sal soluble con aminoácidos y/o proteína parcialmente hidrolizada. |
El término “disponibilidad” ha sido fuente de debate considerable, sin embargo, ha sido definido como la proporción de el mineral/nutriente total en un alimento utilizado para las funciones normales del cuerpo (Fairweather – Trait,1992). Otros han definido biodisponibilidad como la eficiencia con la cual una fuente natural o manufacturada de un elemento distribuye el elemento para almacenar o suplir tejidos metabolicamente activos o para una proteína (Wapnir, 1998). Este también es considerado importante para reflejar la eficiencia de cuales de los nutrientes consumidos son absorbidos del tracto digestivo y así disponibles para almacenamiento o uso (Bender, 1989).
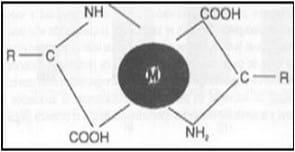 | 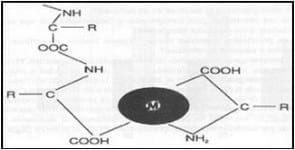 | |
Fig.1 Estructura de un verdadero mineral quelatado. | Fig.2 Estructura básica de los proteinatos minerales. |
Absorción y disponibilidad de los minerales traza orgánicos.
Existen claras diferencias en la biodisponibilidad existente entre fuentes de el mismo metal traza, con metales proteinatos y quelatos, proveyendo fuentes superiores a las fuentes inorgánicas en muchos casos. Durante la digestión, los iones de minerales de las fuentes inorgánicas son liberados y pueden ser recombinados con otros componentes de la digesta en el intestino formando complejos insolubles y por ello excretados, reduciendo su absorción a través del intestino delgado. Esto indica que el grado con el cual estos minerales de la dieta son disponibles para la absorción dependen del grado con el cual ellos forman moléculas complejas en el intestino. En la forma orgánica los elementos traza están protegidos de la acción con otros químicos durante la digestión, haciéndolos más solubles y por lo tanto mejor absorbidos. Son por lo tanto mas biodisponibles y bioactivos y ofrecen al animal ventajas metabólicas que a menudo resultan en un rendimiento mejorado (Close, 1998). Sin embargo un mineral traza puede ser absorbido pero no necesariamente ser utilizado, y su disponibilidad estar siendo baja, aún así los minerales traza utilizando péptidos y/o aminoácidos tiene mayor oportunidad de ser aprovechados por el animal y ser tomados en el intestino (Ashmead et al., 1985; Ashmead, 1993). El mineral dentro del complejo o quelato es una forma químicamente inerte debido a la unión covalente y ligadura iónica por los ligandos aminoácidos, mas estable y menos propenso a las interacciones. El mineral es protegido de factores fisicoquímicos de interacciones negativas con componentes de la dieta tales como el fitato, el cual las lidaguras catiónicas lo hacen indisponible para la digestión (Fair-Trait, 1996). Los minerales traza orgánicos parecen ser eléctricamente neutros en ciertas condiciones de pH. Así, que el quelato mineral es absorbido intacto a la mucosa intestinal, atravesando la membrana de las célula de la mucosa adentro del plasma (Power y Horgan, 2000). Sin embargo, la biodisponibilidad podría ser afectada por varios factores.
Factores que pueden reducir la disponibilidad de los minerales orgánicos
Existen varios factores tales como los factores fisicoquímicos que pueden afectar que sean tomados los nutrientes por el lumen intestinal y éstos sean incorporados en rutas bioquímicas complejas dentro del ambiente celular. Algunos de estos factores están relacionados obviamente a la forma química de el elemento o la presencia de otro iones inorgánicos que compiten por el mismo mecanismo de absorción (Acda and Chae, 2002)(Tabla 3).
Tabla 3. Factores en la dieta que incrementan o disminuyen la absorción, excreción o utilización de ciertos elementos traza.
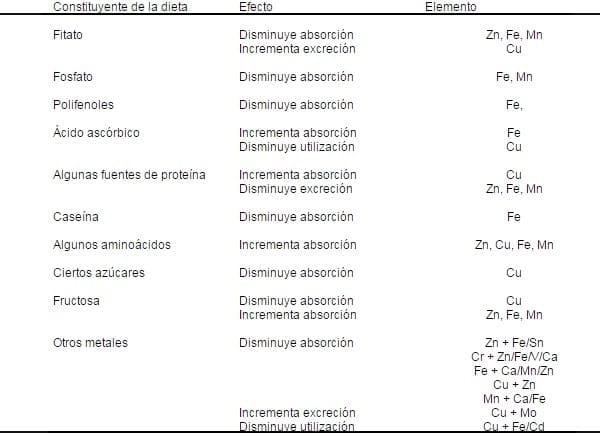 Jonson, 1989 ; Fairweather – Trait, 1992.
Jonson, 1989 ; Fairweather – Trait, 1992.Otros son causados por la interacción de los minerales con los acarreadores de moléculas que mejoran la absorción vía receptores específicos en la mucosa o con otra moléculas orgánicas que la reducen (Acda and Chae, 2002). También se incluyen factores debidos al animal hospedero tales como la edad y especie animal, sexo, etapa de crecimiento, preñez, lactación, estatus nutricional, enfermedades, secreciones gastrointestinales y microflora así como el tiempo de paso gastrointestinal (Johnson, 1989; Fairweather – Trait, 1996).
Aplicaciones de los minerales traza en la nutrición de cerdos
Lo anterior es una revisión breve de las características de los minerales orgánicos, sin embargo en vista del gran número de los minerales traza comerciales disponibles al consumidor, es importante contar con información de los resultados que se han obtenido mediante el uso de los minerales traza orgánicos sobre el comportamiento animal para hacer una selección prudente del elemento a utilizar para que el animal tenga el comportamiento de producción deseado (Acda, 2002). A continuación se reportan algunas investigaciones de gran interés sobre el tema.
Cromo
El cromo es el más comúnmente reconocido por sus funciones fisiológicas y su rol en la potencialización de la acción de la insulina (Mertz, 1993; citado por Money y Cronwell, 1995). El cromo también ha mostrado poseer un rol en el sistema inmune por mejorar el comportamiento del ganado bajo condiciones de estrés (Chang y Mowan, 1992; citado por Money y Cronwell, 1995). Otros roles del Cr incluyen el metabolismo de los ácidos nucleicos (Anderson, 1987; citado por Money y Cronwell, 1995) y lípidos (Mertz, 1993). Una forma del Cr que recientemente ha recibido atención es la fuente orgánica, picolinato de cromo (PCr). Ésta forma consiste de una molécula de Cr3+ complejado con tres moléculas de ácido picolínico. La suplementación con cromo (PCr) en humanos ha demostrado incrementar masa corporal magra y disminuir la grasa del cuerpo (Kaats et al., 1991; citado por Money y Cronwell, 1995). Page et al. (1993) y Lindeman et al. (1995) demostraron respuestas favorables en pruebas hechas en la canal de cerdos con suplementación en la dieta de 200 mg/kg de cromo en la forma de PCR. Sin embargo aunque el PCr ha demostrado mejorar las medidas de la canal en cerdos, éstos efectos sobre el porcentaje de los tejidos de la canal (músculo, grasa, hueso y piel) no se ha reportado ampliamente.
Mooney y Cromwell, (1995) investigaron los efectos del PCr sobre el porcentaje, ganancia total y acumulación de tejidos en la canal y sobre la composición química del músculo en cerdos de crecimiento-finalización. Para ello utilizaron 14 cerdos con un peso inicial promedio de 27 Kg., los cuales fueron alojados individualmente en ambiente controlado con un consumo ad libitum. La dieta basal consistió de maíz y harina de soya para cubrir o exceder los requerimientos principalmente de lisina para cada periodo de crecimiento (NRC, 1988). Además también estuvieron suplementados de vitaminas y minerales establecidos. Los dos tratamientos consistieron de la dieta basal suplementada con 0 (control) o 200 mg/kg de PCr. Éste nivel de fue elegido por lo sugerido por Page et al. (1993), quien con 200 mg/kg mejoró la canal más eficientemente. Las dieta fueron proporcionadas hasta que los animales alcanzaron un peso final de 109± 2 Kg.
Tabla 4. Influencia del picolinato de cromo sobre el comportamiento y medidas de la canal de cerdos crecimiento-finalizaciónabc.
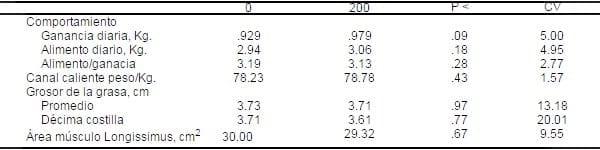 aPromedio de pesos inicial y final fueron 27 y 109 Kg., respectivamente
aPromedio de pesos inicial y final fueron 27 y 109 Kg., respectivamentebMedias de 7 cerdos alojados individualmente
cLos datos de la canal son promedios de ambos lados.
Con la adición de 200 mg/kg PCr tendió (P < .09) a mejorar la tasa de crecimiento, pero no hubo diferencias en consumo de alimento, y no se detectó conversión alimenticia. Mientras que el grosor de la grasa y el área longissimus no fueron influenciados por la suplementación con PCr (Tabla 4).
Tabla 5. Influencia del PCr sobre el porcentaje , ganancia total, y tasas de acreción de tejidos de la canal disectible de cerdos crecimiento-finalizaciónab.
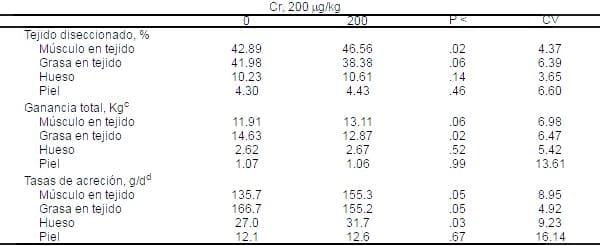 aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza por el peso de la canal caliente.
aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza por el peso de la canal caliente.bLado derecho de la canal.
cPeso final del tejido diseccionado- peso inicial del tejido (basado sobre los cinco cerdos iniciales).
dAcreción total dividida por el número de días en la prueba.
La composición inicial de la canal de cada cerdo fue basado sobre la composición de cinco cerdos que promediaron 27.6 Kg. de peso corporal y 17.2 Kg de peso de la canal (Tabla 5).
La suplementación con PCr incrementó (P<.02) el porcentaje de tejido muscular, disminuyó (P <.06 ) el porcentaje de tejido graso y tendió (P <.14) a incrementar el porcentaje de hueso en la canal diseccionada (Tabla 5). El porcentaje de piel no fue afectada por la adición de PCr. La ganancia total del músculo incrementó por 1.2 Kg. (P<.06) y el tejido graso disminuyó por 1.75 Kg. (P<.02) en cerdos consumiendo CrP, en un 9% de incremento y 12% de reducción. La ganancia total de hueso y piel no fueron afectadas por PCr. Cuando las tasa de acreción diarias fueron determinadas, incrementó de 19.6 g/d en la tasa del músculo (P < .02), y 4.7 g/g en la tasa del hueso (P<.03) ocurrió acreción , con un 11.5 g/d de reducción (P<.05) en la tasa de acreción de grasa. El PCr en la dieta no afectó la tasa de acreción de la piel (Tabla 5).
Los porcentajes de agua y proteína incrementaron en la canal molida en cerdos alimentados con CrP (P<.09 y .004, respectivamente; Tabla 6). El porcentaje de lípidos en la canal molida se redujo (P<.04) en cerdos recibiendo PCr. No hubo cambio en el porcentaje de cenizas en los dos grupos tratados.
Tabla 6. Influencia del PCr sobre la composición química de los tejidos de la canal en cerdos crecimiento-finalizaciónab.
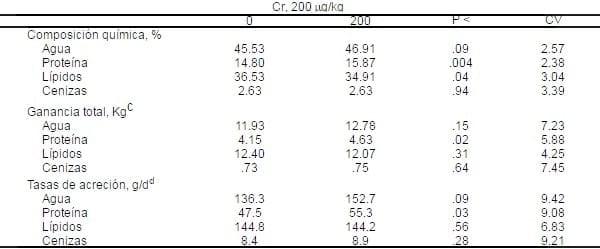 aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza del peso de la canal caliente.
aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza del peso de la canal caliente.bLado derecho de la canal.
cPeso final del tejido diseccionado- peso inicial del tejido (basado sobre los cinco cerdos iniciales).
dAcreción total dividida por el número de días en la prueba.
La ganancia total de proteína en la canal incrementó (P<.02) por 11.5%, y el agua tendió a seguir la misma dirección, en cerdos alimentados con PCr. La ganancia total de lípidos y cenizas en la canal no se afectó por el tratamiento. La suplementación con PCr resultó en un 12 % de incremento (P<.09) en la tasa de acreción diaria de agua y un 16% de incremento (P<.03) en la acreción diaria de proteína en la canal, o un incremento de 16.4 y 7.8 g/d respectivamente. Las tasa de acreción de lípidos y cenizas en la canal no fueron afectadas por el tratamiento (Tabal 6).
Tabla 7. Influencia del PCr sobre la composición química del tejido muscular diseccionado en cerdos crecimiento-finalizaciónab.
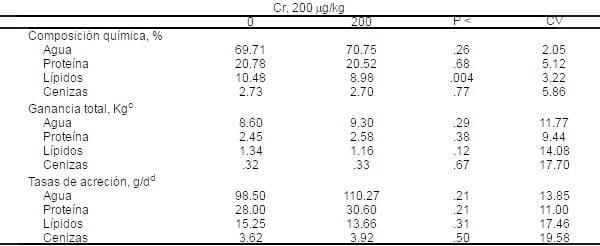 aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza de el peso de la canal caliente.
aMedias de siete cerdos alojados individualmente por tratamiento. Todas las medias son ajustadas por covarianza de el peso de la canal caliente.bLado derecho de la canal.
cPeso final del tejido diseccionado- peso inicial del tejido (basado sobre los cinco cerdos iniciales).
dAcreción total dividida por el número de días en la prueba.
Los cerdos consumiendo el PCr en la dieta tuvieron un porcentaje más bajo (P<.004) de lípidos en el músculo diseccionado (Tabla 7), pero los porcentajes de agua, proteína y cenizas en el músculo fueron similares entre tratamientos. La suplementación de PCr en la dieta también tendió (P<.12) a disminuir la ganancia total de lípidos, pero esto tuvo efecto no significativo sobre la ganancia total de agua, proteína o ceniza. La tasa de acreción de agua y proteína en el músculo diseccionado incrementó numéricamente en cerdos alimentados con PCr, pero éstos incrementos no tuvieron significancia (P=.21). La tasa de acreción de lípidos en el músculo diseccionado disminuyó por 1.59 g/d debido al Cr en la dieta, pero ésta diferencia no fue significativa (P=0.31)
Previos experimentos han demostrado resultados variables en la tasa de crecimiento y relación alimento:ganancia en cerdos alimentados con dietas suplementados con PCr.Page et al. (1993) reportó un incremento en la tasa de crecimiento en un experimento pero en otros no cambió la eficiencia alimenticia. Lindemann et al. (1995) no observaron cambio en la tasa de crecimiento pero encontraron una mejora en la relación alimento:ganancia con la adición de Cr en forma de PCr a 200, 250, o 500 mg. En la presente investigación (Mooney y Cromwell, 1995), 200 mg/Kg. de cromo incrementó (P<.09) la tasa de crecimiento pero no hubo mejora en la eficiencia alimenticia. La carencia de mejoramiento en la relaciónalimento:ganancia estuvo asociada con el incremento numérico en el consumo de alimento.
El efecto de 200 mg/Kg. de Cr como PCr en las medidas de la canal de cerdos fue reportado por Page et al. (1993). Ellos reportaron que en promedio incrementó de 15.3% en el área del músculo longissiums y en promedio disminuyó de 17.3% en la 10ª costilla. Sin embargo, experimentos mas recientes no han demostrado mejoramientos consistentes en las medidas de las canal. Lindemann et al. (1995) no observó cambios en el área del lomo o el área longissimus en cerdos alimentados con dietas suplementadas con 250 o500 mg de Cr como PCr de 40.9 a 98.8 Kg. de peso vivo (PV). Los resultados de este experimento no tuvieron mejoras significativas en el lomo, décima costilla o área longissimus cuando los cerdos fueron alimentados con PCr. El presente experimento está de acuerdo con Lindemann et al. (1995) donde tampoco se obtuvieron diferencias significativas en las medidas de el lomo, décima costilla o área longissimus cuando los cerdos fueron alimentados con PCr.
En cuanto al mejoramiento del porcentaje magro en la canal involucrando suplementación con PCr, Page et al. (1993) encontró un 2.95 de incremento en unidades porcentuales (P<.01) mientras que Lindemann et al. (1995) mostraron unas 3.0 unidades porcentuales de incremento (P<.04) en el porcentaje de músculos en cerdos tratados con PCr. Sin embargo no se tuvieron diferencias en las medidas de la canal, los procedimientos de disección usados en el presente experimento revelaron que un aumento de 3.7% en el músculo diseccionado (P<.02) y un 3.6 % disminuyó en el porcentaje de grasa diseccionada (P<.06).
La composición de la canal hubo un aumento de agua (P<.09) y proteína (P<.004) y una disminución de lípidos (P<.12) esto concuerda con lo reportado con Boleman et al. (1995).
La investigación indica que la suplementación con PCr incrementó el porcentaje, ganancia total y la tasa de acreción del músculo y disminuye el porcentaje de ganancia total, y acreción de grasa en cerdos en la etapa crecimiento-finalización.
Más estudios demuestran que el Cr también mejora el comportamiento reproductivo (tiene efectos sobre hormonas como la oxcitocina y progesterona; aumenta el número de lechones por camada y destetados; aumenta la tasa de fertilidad; hay menos abortos y momificaciones; aumenta la tasa de concepción; aumenta la viabilidad de embriones y también aumenta la sensibilidad de los tejidos a la insulina, mejorando la actividad reproductiva de las hembras (Lindemann, 1995 ab; Evans, 1992).
Selenio
Olson et al. (1970) determinó que la seleniometionina fue el principal aminoácido seleno natural en trigo, y considerado ser la principal forma de selenio (Se) encontrado en la mayoría de los granos que no son almacenados (Muth and Olfield, 1967; citado por Mahan et al., 1999). El perfil de análogos de Se fueron encontrados en levaduras enriquecidas con Se (por tener 40% de selenometionina, 15% de selenocistina, y un menor porcentaje de otros aminoácidos análogos) (Kelly and Power, 1995; citado por Mahan et al. (1999). Cuando la levadura fuente de selenio orgánico fue administrada a cerdos de crecimiento y finalización, hubo una mayor deposición de Se en el músculo que cuando el elemento proveído era una fuente inorgánica Mahan y Parret, (1996). Aunque la deposición de Se en el tejido del cerdo fue mayor cuando la forma orgánica del Se fue proporcionada, el selenito de sodio fue mas efectivo en alcanzar la actividad máxima de la glutation peroxidasa (GSH-Px).
Ambos el Se y la vitamina E se han reconocido por tener propiedades antioxidantes, pero el selenito de sodio también puede actuar como prooxidante, particularmente en altos niveles en la dieta (Seko et al., 1989; citado por Mahan et al. 1999), mientras que la seleniometionina no posee ésta propiedad (Spallholz, 1994).
Mahan et al. (1999) evaluaron la eficiencia de dos fuentes de Se (selenito de sodio o levadura enriquecida con Se) para cerdos crecimiento-finalización. Evaluaron el comportamiento del cerdo, la actividad en suero de GSH-Px, respuestas del tejido y la canal y las medidas de calidad del lomo.
En el primer factor se evaluaron los efectos del selenito de sodio (inorgánico) o la levadura enriquecida con Se (orgánico) como dos fuentes de Se en la dieta. Cuatro niveles de Se fueron agregados en la dieta, .05, .10, .20, o .30 mg Se/Kg de dieta constituida con el segundo factor, un grupo control con la adición de Se no fortificado en la dieta basal. La dieta basal constituyó de maíz, harina de soya y lisina para formular .80% y .70% de lisina para lo períodos de crecimiento y finalización. Fueron agregadas vitaminas y minerales, con excepción del Se, para cubrir o exceder los requerimientos según NRC (1988), el consumo fue ad libitum.
Los efectos de la fuente de Se y los niveles de Se en la dieta sobre las respuestas de comportamiento del cerdo son presentados en la Tabla 8. La concentración de la dieta basal estuvo en un rango de .05 a .08 mg Se/Kg de dieta. Los resultados de comportamiento de los cerdos durante el crecimiento, finalización y el total del periodo experimental no mostraron tener algún efecto (P>.15) con la fuente de Se o nivel de Se sobre ganancia de peso en cerdo, consumo de alimento o la relación ganacia:alimento. Estos resultados coinciden con los de Thomke et al. (1965), Wastell et al. (1972), Ku et al., (1973), y Mahan y Parret, (1996), quienes también demostraron que no hubo respuestas en la tasa de crecimiento cuando el Se fue en forma orgánica o inorgánica y fue agregado en varios niveles para dietas basadas en cereales en dietas para cerdos crecimiento-finalización (Tabla 8).
Tabla 8. Efectos de las fuentes de Se y niveles sobre el comportamiento en cerdos crecimiento-finalización.
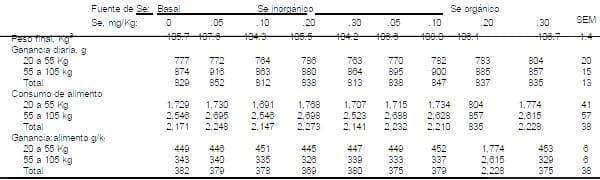 aCada media representa 39 cerdos por tratamiento con peso corporal inicial de 20.4 Kg.
aCada media representa 39 cerdos por tratamiento con peso corporal inicial de 20.4 Kg.Tabla 9. Efectos de las fuentes de Se y niveles sobre el comportamiento en cerdos crecimiento-finalización.
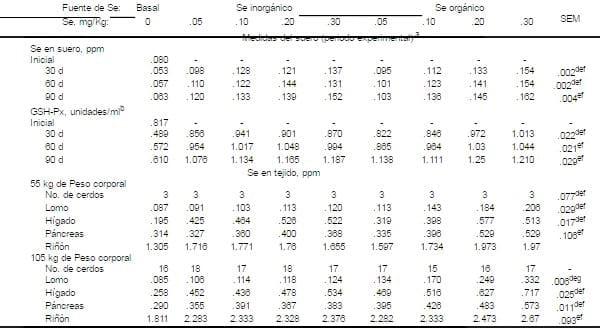 a Cinco cerdos fueron sangrados
a Cinco cerdos fueron sangrados b Una es igual a la actividad 1 m m de NADPH oxidada por minuto/por mililitro se suero
c El peso corporal promedio de los cerdos nuertos fue de 55 Kg.
d Respuesta de interacción en la fuente de Se en la dieta x el nivel de Se (P<.01).
e Respuesta cuadrática para el nivel de selenito de sodio (P<.01). La regresión incluye la comparación de la dieta basal.
f Respuesta cuadrática para el nivel de levadura enriquecida con Se (P<.01). La regresión incluye la comparación de la dieta basal.
g Respuesta lineal para el nivel de levadura enriquecida con Se (P<.01). La regresión incluye la comparación de la dieta basal.
En cuanto a las medidas de suero , cuando los cerdos fueron alimentados con ambas dietas basales , la concentración de Se y la actividad de la GSH-Px en el suero los valores disminuyeron al inicio del experimento (día 30) del periodo de colección (Tabla 8). Aunque las concentraciones de suero fueron más bajas después en el día 30 de alimentación, éstas incrementaron en cada medida de periodos subsecuentes, aún cuando los cerdos en crecimiento fueron alimentados con la dieta basal. Debido a que los cerdos de mayor edad tenían mayor consumo de alimento, esto podría explicarse las mayores concentraciones de Se en suero aún cuando la dieta tenía el Se no fortificado.
Las concentraciones de suero incrementaron (P<.01) conforme los niveles de Se en la dieta incrementaron en cada período. Hubo sin embargo, una interacción nivel de selenio y fuente de selenio (P<.01) para el período de el día 30 y el día 60. El Se en suero fue un poco más alto .05 mg/Kg en los niveles de selenio, cuando el selenito de sodio fue la fuente de Se, pero, como el nivel de selenio en la dieta incrementó, las concentraciones de Se en suero fueron mayores cuando el Se orgánico fue la fuente de Se proveída.
La actividad de la GSH-Px en suero se incrementó de los 30 a los 90 días por periodo de muesteo cuando se proporció la dieta basal. Ésta respuesta fue consistente con las incrementadas concentraciones de Se en suero en cada período medido en éste grupo tratado. La actividad de la GSH-Px incrementó cuadraticamente (P<.01) conforme los niveles de Se incrementaron para ambas fuentes de Se. Sin embargo, las actividades mas bajas en el período 30 y 60 fueron de .05 mg/kg cuando la dieta proveyó Se orgánico comparada con el Se inorgánico, la interacción fue significativa (P<.01) solamente para el período 30 est concuerda con los datos de Mahan y Parrett, (1996)(Tabla 9).
Cu
El cromo promueve el crecimiento cuando se alimenta de 100 a 250 mg/Kg (Bunch et al., 1965), aunque éste método de acción es aún incierto. Cromwell et al. (1989) y Bowland(1990), sugirieron sin embargo, que la estimulación del crecimiento por cobre estuvo relacionada a la solubilidad de la fuente de cobre; por eso el cobre es más disponible para la absorción. Wapnir y Balkman, 1990 sugirió que el cobre unido a péptidos y aminoácidos en la digesta, ayudan la absorción del cobre. Zhouet et al.(1994b) y Coffey et al. (1994) reportaron un incremento en el comportamiento de cerdos en crecimiento alimentados con un complejo sobre la lisina (CLys) comparado con cerdos alimentados con sulfato de cobre (CuSO4). Sin embargo en otros ensayos usando CuLys (Coffey et al., 1994), metionina de cobre (Bunch et al., 1965), o complejo de cobre (Stansbury et al., 1990) no tuvieron diferencias entre el complejo éstas dos fuentes. Otros investigadores también han reportado, que no hay diferencias en la actividad de promover el crecimiento entre las dos fuentes de cobre (Apgar et al., 1995). Aoyagi y Baker (1994) reportaron que los complejos CuLys y cobre-metionina que se les proporcionó a pollos no tuvieron una menor disponibilidad pero cuando se les alimentó en combinación con elevados niveles de L-cistina y L-ácido ascórbico, los dos componentes de la dieta sí tuvieron una menor absorción de Cu. Por lo tanto éstas respuestas sugieren que el Cu en formas orgánicas puede ser absorbido de diferente manera y afectado por otros aditivos en la dieta.
Apgar y Kornegay, (1996) compararon el balance mineral y digestibilidad de la materia seca en cerdos de finalización alimentados con altos niveles de cobre en CuSo4 o el complejo CuLys en la presencia de cantidades excesivas (tres a cuatros veces los niveles recomendados) de otros minerales traza (Fe, Cu, Mn y I). Los coeficientes de absorción también fueron comparados usando el óxido de cromo como método indicador. Se utilizaron 24 cerdos con un peso inicial promedio de 83.7 kg en la Prueba 1 y 80.6 Kg. en la Prueba 2. Los tratamientos en la dieta fueron 1) una dieta basal basada en harina de soya –maíz con 16% de proteína (36 mg/Kg de Cu), 2) fue la dieta basal + 200 mg/Kg deCu de CuSO4 y ·5H2O y 3) La dieta basal + 200 mg/Kg de Cu de CuPlex 100. El óxido crómico fue agrgado en un nivel de .05% en una mezcla. La absorción también fue determinada por la colección total del método.
La ganancia diaria promedio tendió a ser mejor para cerdos alimentados con CuLys que para lo cerdos alimentados con CuSo4 (Tabla W, P<.02). Los cerdos del grupo control tuvieron lo valores intermedios. Las concentraciones de Cu en heces fue mayor (P<.001) para los cerdos alimentados con Cu elevado de ambas fuentes que los del grupo control. Los cerdos alimentados con Cu de CuLys tendieron (P<.09) a tener mayores concentraciones de Cu fecales que los cerdos alimentados con el cu de CuSo4. los porcentajes de Fe y Znfueron similares entre grupos de tratamientos, pero las concentraciones fecales de Cr tendieron a ser mayores (P<.10) en cerdos alimentados con el Cu de CuLys que los cerdos alimentados con Cu de CuSO4; el control fue intermedio.
Tabla 10. Ganancia diaria y porcentaje de concentración mineral en alimentos y heces de cerdos en finalización alimentados con 0 o 200 mg/Kg de Cu agregado en la dieta ya sea CuSO4 o el complejo CuLys (Prueba 1 y 2 combinado)a
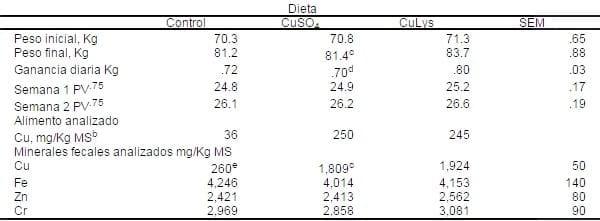 aLos datos son medias de 16 observaciones (2 diferentes pruebas, cuatro cerdos por tratamiento por ensayo, y dos periodos de colleción por ensayo).
aLos datos son medias de 16 observaciones (2 diferentes pruebas, cuatro cerdos por tratamiento por ensayo, y dos periodos de colleción por ensayo).bLas concentraciones de Fe, Zn y Cr (mg/Kg de MS) fueron de 530, 36%, y 380, respectivamente.
cCuSO4 diferente de CuLys (P<.09).
dCuSO4 diferente de CuLys (P<.02).
eLos cerdos control diferentes de cerdos con CuSO4 y CuLys (P<.09).
Para el consumo de materia seca (MS), excreción de MS en heces, la cantidad absoluta de materia seca digerida y la digestibilidad no difirió entre tratamientos en los grupos tratados (Tabla 11).
Tabla 11. Materia seca y absorción de Cu y Cr datos del balance en cerdos en finalización alimentados con 0 o 200 mg/Kg de Cu con CuSO4 o el complejo CuLys (Prueba 1 y 2 combinado)a
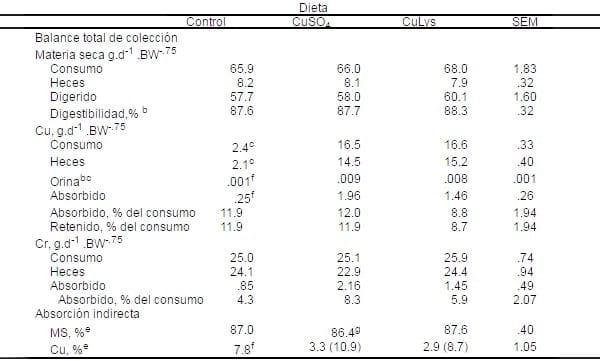 aLos datos son medias de 16 observaciones (2 diferentes pruebas, cuatro cerdos por tratamiento por ensayo, y dos periodos de colleción por ensayo).
aLos datos son medias de 16 observaciones (2 diferentes pruebas, cuatro cerdos por tratamiento por ensayo, y dos periodos de colleción por ensayo).bInteracción prueba x tratamiento (P<.05).
cEfecto de la semana (P<.05).
dInteracción prueba x tratamiento (P<.01).
eInteracción prueba x tratamiento (P<.001).
fCmedias para los controles, fueron diferentes de las medias para CuSO4 y CuLys (P<.001).
gCuSO4 menor que CuLys (P<.04).
hEl valor en paréntesis fue calculado después de ajustar la concentración de Cr por el Cr recobrado.
El consumo de Cu y excreción de Cu en heces y orina fue mejor (P<.001) para cerdos alimentados con Cu elevado en ambas fuentes que para los controles, no hubo diferencia entre las dos fuentes. La excreción de Cu en la orina fue afectado por la semana (P<.05); los valores fueron mejores para la semana 2 que para la 1 (.0075 contra .0045, respectivamente). La cantidad absoluta de cobre absorbido fue mayor para cerdos alimentados con Cu de las fuentes CuSO4 o CuLys (P<.001) que para los controles. Sin embargo, el porcentaje de cu absorbido y retenido no difirió entre grupos tratados.
El consumo de Cr y excreción de Cr en heces no difirieron entre grupos tratados. Una interacción entre prueba y tratamiento (P<.05) fue observado para la excreción en heces de Cr: en Prueba 1 la alimentación de los cerdos con Cu de la forma CuSO4 tuvieron los valores más altos en heces. En la Prueba 2 los cerdos alimentados con Cu de la forma CuLystuvieron los valores más bajos en heces.
El porcentaje de cromo absorbido no difirió entre tratamientos. Se observó una interacción entre prueba y tratamiento (P<.01) fue observado para la excreción en heces de Cr: en Prueba 1 la alimentación de los cerdos con Cu de la forma CuSO4 tuvieron los valores más altos en heces. En la Prueba 2 los cerdos alimentados con Cu de la forma CuLys tuvieron los valores más bajos en heces.
Los niveles de Cu absorbido estimularon más el crecimiento comparado con el grupo control de cerdos, pero no se observaron diferencias entre las fuentes proporcionadas de Se la forma orgánica y la inorgánica.
Referencias
- AAFCO. 1998. Officials Publication of the Association of American Feed Control Officials Incorporated (Paul. M. Bachman, ed). Page 237-238.
- Acda and Chae, 2002
- Agricultural Research Council (ARC). 1981. The Nutrient Requirements of Pigs. Commonwealth Agricultural Bureaux, Slough.
- Ammerman, C.B., Henry, P.R. y Miles, R.D. (1998) Supplemental organically bound mineral compounds in livestock nutrition. En: Recent Advances in Animal Nutrition. Ed. Garnsworthy, P.C. y J. Wiseman. pp: 67-91. Nottingham University Press, Nottingham, Reino Unido.
- Anderson, 1987; citado por Money y Cronwell, 1995
- Ashmead, H. D., D. J. Graff and H. H. Ashmead, 1985. Intestinal absorption of metal ions and chelates. C. C. Thomas, Springfield, Illions.
- Ashmead, H. D., 1993. Comparative intestinal absortion and subsequent metabolism of metal amino acid chelates and inorganic metal salts. In: The roles of amino acid chelates in animal nutrition. Noyes Publishers, New Jersey, 306-319.
- Baker, D.H. y Johnson, B. (1997) Feedstuffs 69 (19): 14.
- Bender, A.E. 1989. Nutritional significance of bioavailability. In: Nutrient Availability: Chemical and Biological aspect. (D.A.T. Southgate, I.T. Johnson and G.R. Fenwik, eds). Special Publication Nho. 72Royal Society of Chemistry, Cambridge, pp 3-9.
- Boleman et al. (1995).
- Close, 2000
- Close, 2002.
- Close, W. H., 1998. The role of trace mineral proteinates in pig nutrition. In: Biotechnology in the Feed Industry. (T. P. Lyos and K. A. Jacques, eds.). Nottingham University Press.Nottingham, UK., pp: 469-483.
- Close, W. H., 1999. Organic minerals for pigs: an update. In: Biotechnology in the Feed Industry. (T. P. Lyos and K. A. Jacques, eds.). Nottingham University Press. Nottingham, UK., pp: 51-60.
- Cole, J.A. (1991) Pig News & Information 12 (3): 393-401.
- Chang y Mowan, 1992; citado por Money y Cronwell, 1995
- Fairweather – Trait, S. J., 1996. Bioavailability of dietary minerals. Bioch. Soc. Trans., 24: 775-780.
- Fairweather-Trait, S.J. 1992. Bioavailability of trace elements. Food Chem.43: 213-217.
- Johnson, P. E. 1994. Trace Minerals and Fertility in Dairy Cattle. Biotechnology in the Feed Industry. Proceedings of the 10th – 15th Symposium 1994 – 1999. Nottingham University Press. England.
- Jonson, P. E., 1989. What can in vitro methods tell us about mineral availability Biological Trace Minerals Research, 19: 3-10.
- Kaats et al., 1991; citado por Money y Cronwell, 1995
- Kelly and Power, 1995; citado por Mahan et al. 1999
- Lindemann, 1995 ab; Evans, 1992)
- Lindemann, M. 2001. Revisión del Valor del Cromo orgánico en cerdos. Biotecnología en la industria de la Alimentación Animal. Alltech de México S. A. de C. V. México.
- Lindemann, M. D. 1998. Organic Chromium – an Exciting Beginning, a Promising Future. Biotechnology in the Feed Industry. Proceedings of the 10th – 15th Symposium 1994 – 1999. Nottingham University Press. England.
- Mahan. D. C. 1995. Selenium Metabolism in Animals: What Role does Selenium Yeast Have? Biotechnology in the Feed Industry. Proceedings of the 10th – 15th Symposium 1994 – 1999. Nottingham University Press. England.
- Mahan. D. C. 1994. Organic Selenium Sources for Swine, How do They Compare with Inorganic Selenium Sources? Biotechnology in the Feed Industry. Proceedings of the 10th – 15thSymposium 1994 – 1999. Nottingham University Press. England.
- Mahan et al. (1999)
- Mahan y Parret, 1996
- Mertz, 1993; citado por Money y Cronwell, 1995
- Mooney y Cromwell, (1995)
- Muth and Olfield, 1967; citado por Mahan et al., 1999
- National Research Council. 1988. Nutrient Requirements of Swine. 9th Revised Edition. National academy Press, Washington.
- National Research Council. 1998. Nutrient Requirements of Swine. 10th Revised Edition. National academy Press, Washington.
- Olson et al. (1970)
- Power y Horgan, 2000. Alltech 2000.
- Power, R and K. Horgan, 2000. Biological chemistry and absortion of inorganic and organic trace metals. In: Biotechnology in the Feed Industry. (T. P. Lyos and K. A. Jacques, eds.). Nottingham University Press. Nottingham, UK., pp: 277-291.
- Seko et al., 1989; citado por Mahan et al. 1999
- Spallholz, 1994
- Spears, J.W. (1989) J. Anim. Sci. 67: 835-843
- Wapnir, 1998
- Wapnir, R.A. 1998. Cooper absorption and bioavailability. A.M. J . Clin. Nutr. 67(Suppl):1054S-1060S.
Temas relacionados
Autores:

Universidad Autónoma de Chihuahua - Mexico
Recomendar
Comentar
Compartir

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.




Te puede interesar

Yara Animal Nutrition