Necesidades Proteicas en el Ganado Vacuno Lechero
Publicado: 29 de noviembre de 2006
Por: Andrés L. Martínez Marín, Veterinario
Las proteínas son compuestos nitrogenados (contenido medio en nitrógeno de 16%) integrantes fundamentales de los tejidos animales. Las proteínas representan una proporción variable del peso vivo del animal (10-20%) pero sin embargo el contenido en la masa corporal desengrasada es prácticamente constante (21%). La mayoría de las proteínas forman parte de la estructura de tejidos y órganos, no existiendo un tejido específico para el almacenamiento de la proteína excedentaria. Una pequeña fracción de la proteína corporal tiene un papel funcional en el organismo en forma de enzimas, hormonas, etc. Las proteínas están constituidas por moléculas más sencillas denominadas aminoácidos, cuyo número y distribución en cada proteína está codificado genéticamente. El número de aminoácidos es de veinte, de los cuales diez son esenciales porque la biosíntesis es insuficiente (arginina, histidina, isoleucina, leucina, metionina, fenilalanina, triptófano y valina) o imposible (lisina y treonina), por tanto su demanda debe cubrirse con aportes en la dieta. Los restantes diez aminoácidos (glicina, alanina, serina, tirosina, ácido aspártico, asparagina, ácido glutámico, glutamina, prolina y cistina) son no esenciales ya que pueden ser sintetizados desde compuestos intermediarios del metabolismo hidrocarbonado o desde otros aminoácidos. A diferencia de los monogástricos, en los rumiantes las necesidades de aminoácidos son cubiertas parcialmente a través de la proteína microbiana sintetizada en rumen ya que la flora ruminal es capaz de sintetizar todos los aminoácidos, incluyendo los esenciales, incluso desde compuestos nitrogenados sencillos.
Los animales sufren pérdidas de nitrógeno con las heces, la orina, a través de la piel y con la leche. Por otro lado, la masa de proteína corporal aumenta en los animales jóvenes en crecimiento y en las hembras en gestación. En todos los casos se generan unas necesidades de aminoácidos determinadas, al ser estos compuestos la única forma nitrogenada utilizable por el organismo animal.
LAS NECESIDADES PROTEICAS DEL GANADO VACUNO LECHERO.
La principal diferencia entre los rumiantes y los animales monogástricos se refiere a los procesos digestivos, particularmente a la relación simbiótica establecida entre el rumiante y la microflora existente en el rumen e intestino grueso. Más del 60% de la energía (en forma de ácidos grasos volátiles) y del 50% de la proteína utilizada por el animal rumiante tiene su origen en la digestión microbiana de los alimentos en el rumen. La digestión que ocurre en intestino delgado es enzimática y similar a la de los animales monogástricos. En intestino grueso existe fermentación microbiana de los restos alimenticios no digeridos en las porciones anteriores del tubo digestivo pero aunque esto supone un aporte de energía importante, en torno a un 15% , la proteína aquí sintetizada no puede ser absorbida y se pierde en las heces.
La complejidad del proceso digestivo de los rumiantes es determinante a la hora de intentar valorar las necesidades y los aportes de proteína realmente utilizables por el organismo. Los compuestos nitrogenados presentes en los alimentos (proteína verdadera y compuestos nitrogenados no proteicos), son utilizados por los microorganismos del rumen para la síntesis de compuestos nitrogenados microbianos, principalmente proteína. Las células microbianas (mayoritariamente bacterias y protozoos) son arrastradas junto a partículas alimenticias no fermentadas y células epiteliales descamadas hacia las porciones posteriores del tramo digestivo donde ocurre digestión enzimática y absorción de los diferentes nutrientes. El conjunto de aminoácidos disponibles para su absorción en intestino (ver más adelante) constituye la denominada proteína "metabolizable" o "absorbible" que puede ser realmente utilizada por el organismo.
La complejidad del proceso digestivo de los rumiantes es determinante a la hora de intentar valorar las necesidades y los aportes de proteína realmente utilizables por el organismo. Los compuestos nitrogenados presentes en los alimentos (proteína verdadera y compuestos nitrogenados no proteicos), son utilizados por los microorganismos del rumen para la síntesis de compuestos nitrogenados microbianos, principalmente proteína. Las células microbianas (mayoritariamente bacterias y protozoos) son arrastradas junto a partículas alimenticias no fermentadas y células epiteliales descamadas hacia las porciones posteriores del tramo digestivo donde ocurre digestión enzimática y absorción de los diferentes nutrientes. El conjunto de aminoácidos disponibles para su absorción en intestino (ver más adelante) constituye la denominada proteína "metabolizable" o "absorbible" que puede ser realmente utilizada por el organismo.
La utilización de la proteína metabolizable es diferente para cada proceso productivo en función principalmente de su composición en aminoácidos. La conversión de la proteína metabolizable en tejidos o proteína láctea conlleva unas pérdidas asociadas de nitrógeno en heces y orina debidas a los gastos inevitables de síntesis y a los desequilibrios entre los aminoácidos disponibles. Considerando esto, las necesidades de proteína neta corresponden a la proteína retenida realmente por el organismo para el mantenimiento, el crecimiento, la lactación o la gestación, y la proteína metabolizable es la suma de dichas necesidades proteicas y las pérdidas asociadas a su síntesis.
En esta primera exposición nos vamos a centrar en la estimación de las necesidades de proteína (y aminoácidos) y en la eficiencia de utilización para cada una de las funciones corporales.
DINAMICA DEL NITROGENO EN EL ORGANISMO.
- Absorción de compuestos nitrogenados (FIGURA I).
DINAMICA DE LOS COMPUESTOS NITROGENADOS EN EL ORGANISMO.
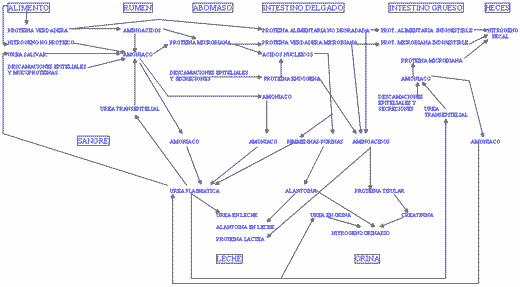
Al intestino delgado llega proteína microbiana (proteína verdadera, paredes celulares y ácidos nucleicos), proteína alimentaria no degradada, proteínas de origen endógeno (células de descamación y secreciones) y amoníaco. La digestión enzimática es similar a los monogástricos con algunas diferencias:
- La cantidad de nitrógeno endógeno es importante en relación con el nitrógeno microbiano y alimentario.
- El ritmo de neutralización duodenal de la digesta es inferior a los monogástricos, lo que retrasa la actuación de las enzimas intestinales pero permite que la proteolisis debida a la pepsina abomasal se mantenga en duodeno.
- El punto de máxima actividad de las proteasas pancreáticas se retrasa hasta el yeyuno medio, situándose aquí el punto de mayor absorción de aminoácidos.
La proteína verdadera microbiana constituye aproximadamente el 85% de la proteína microbiana y un 75% es digestible. La digestibilidad media de la proteína alimentaria no degradada en rumen es de un 85%. Ambas fuentes proveen, en proporciones variables según el tipo de ración, el conjunto de aminoácidos disponibles para su absorción intestinal. La fracción no digestible se elimina con las heces. La absorción a través de la pared intestinal es diferencial, de forma que de media el 60% de los aminoácidos esenciales que atraviesan el píloro y el 43% de los no esenciales son absorbidos.
La alta actividad de las nucleasas del jugo pancreático en los rumiantes se justifica por el elevado contenido en ácidos nucleicos de las células microbianas (más del 15% de nitrógeno microbiano), y permite la liberación de fósforo que puede ser utilizado por el organismo. Por el contrario, los restantes productos liberados en la digestión de los ácidos nucleicos (purinas y pirimidinas) son inutilizables y contribuyen a los gastos de excreción.
El amoníaco libre absorbido proviene de la degradación microbiana ruminal de los aminoácidos y los compuestos nitrogenados no proteicos y representa el exceso no utilizado para la síntesis de proteína. La absorción ocurre a través del epitelio ruminal, desde intestino delgado y también desde intestino grueso. La cantidad de nitrógeno absorbido como amoníaco es variable según el tipo de ración consumida pero puede alcanzar valores muy elevados en raciones ricas en nitrógeno no proteico.
- Metabolismo tisular.
Los aminoácidos absorbidos y los endógenos procedentes del proceso de renovación tisular (ver más adelante) constituyen un conjunto de aminoácidos libres que pueden utilizarse para procesos de síntesis de proteína, como precursores glucogénicos u oxidarse completamente para generar energía. La contribución de los aminoácidos al metabolismo energético corporal es muy importante en los rumiantes debido a la baja tasa de absorción de glucosa con relación a las necesidades totales diarias, sobre todo del tejido nervioso y de la ubre.
El metabolismo visceral y hepático es el determinante principal de la cantidad y proporciones de aminoácidos disponibles en la sangre (absorbidos y endógenos). (FIGURA II). En los tejidos digestivos ocurre una intensa utilización de aminoácidos para la renovación celular y la provisión de energía. Debido a que la mayor proporción de tejidos no participa en la absorción de aminoácidos, se estima que en el conjunto de las vísceras drenadas por la vena porta en torno al 78% de los aminoácidos utilizados es de origen endógeno y el resto procede de los aminoácidos absorbidos de la luz intestinal.
Figura II / FLUJO DE LOS AMINOACIDOS EN EL ORGANISMO.
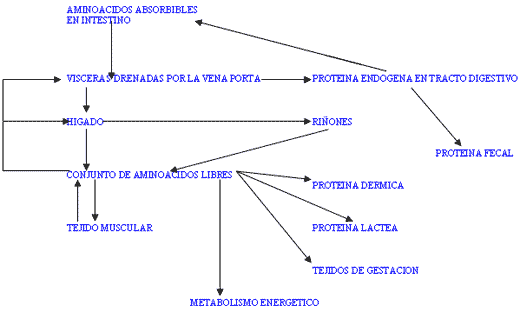
Figura II / FLUJO DE LOS AMINOACIDOS EN EL ORGANISMO.
En hígado ocurre catabolismo parcial (desaminación) de aminoácidos, absorbidos y endógenos para la síntesis de glucosa, de forma particularmente intensa durante períodos de subnutrición energética o de elevada demanda de glucosa (periparto y comienzo de la lactación), oxidación completa para generar energía, o conversión de unos aminoácidos en otros y síntesis de aminoácidos no esenciales desde oxoácidos (transaminación).
La ubre utiliza los aminoácidos extraídos de la sangre para la síntesis de proteína láctea y como fuente de energía o de glucosa. Los tejidos de gestación utilizan los aminoácidos absorbidos para la síntesis proteica y como fuente de energía por oxidación. Los riñones extraen aminoácidos posiblemente para su conversión a glucosa a la par que sus metabolitos contribuyen al control del equilibrio ácido-básico. El tejido muscular es el principal proveedor de aminoácidos endógenos a los restantes tejidos: glutamina como combustible para las vísceras del sistema porta y los riñones, y alanina como precursor glucogénico al hígado.
Los procesos de síntesis de proteína incluyen el crecimiento de la masa muscular en animales jóvenes, la síntesis de proteína de la leche, el crecimiento de los tejidos fetales y anejos, el desarrollo de la ubre y el mantenimiento de la integridad corporal (síntesis vs. degradación).
En conjunto existe una estrecha relación entre el metabolismo proteico de los diferentes tejidos (FIGURA II).
3) Metabolismo de la urea y otros compuestos nitrogenados.
El exceso de nitrógeno presente en el organismo debe ser detoxificado y eliminado. La principal vía de eliminación de los compuestos de detoxificación es la orina.
El amoniaco absorbido desde el tracto intestinal o generado en el catabolismo tisular de los aminoácidos y en la degradación de las pirimidinas, debe ser detoxificado mediante su conversión a urea, siendo éste el principal mecanismo de eliminación del nitrógeno en exceso presente en el organismo. Una fracción es detoxificada en forma de glutamina que actúa como transportador del amoníaco a los riñones donde es eliminado tal cual. También, parte del amoníaco libre es utilizado en las reacciones de transaminación para la síntesis de aminoácidos no esenciales desde los cetoácidos correspondientes. La síntesis de urea ocurre a través del denominado ciclo de la ornitina (FIGURA III). En este proceso, los dos grupos amino necesarios son cedidos por el ácido glutámico, el primero directamente por desaminación oxidativa, y el segundo por transaminación del oxalacetato para rendir ácido aspártico, la ornitina necesaria procede de la arginina, se consume aspartato y se genera fumarato, la energía necesaria procede del adenosintrifosfato (ATP). Existe una estrecha relación entre este ciclo y el metabolismo energético (ver más abajo).
Figura III / RELACION ENTRE LA EXCRECIÓN DE AMONIACO Y EL METABOLISMO ENERGETICO
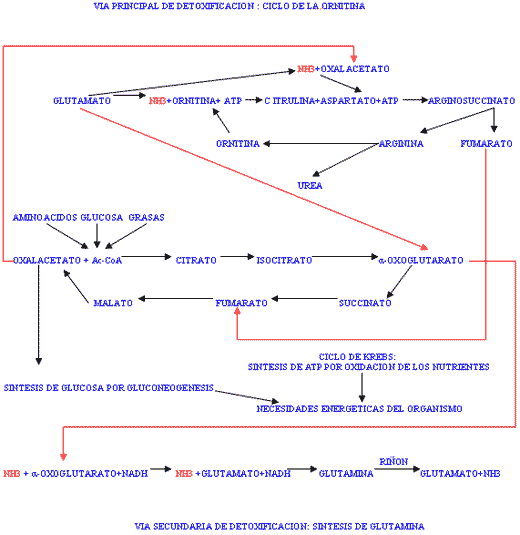
Figura III / RELACION ENTRE LA EXCRECIÓN DE AMONIACO Y EL METABOLISMO ENERGETICO
Otros compuestos que sirven como vehículos para la eliminación de nitrógeno son la alantoína y el ácido úrico, originados en el catabolismo de las purinas, la creatinina y la metil-histidina, que se relacionan con la renovación de la masa muscular, y los ácidos hipúrico y fenacetúrico que son conjugados de glicina con metabolitos microbianos.
Los ácidos nucleicos endógenos contribuyen mínimamente al conjunto de productos nitrogenados de desecho debido a que son estrictamente reciclados. Se calcula que un 90% de las purinas liberadas en la degradación de los ácidos nucleicos son reutilizadas.
La urea presente en el plasma puede seguir varios caminos:
a) Reciclado ruminal: si existe un déficit de nitrógeno para el crecimiento microbiano, el organismo es capaz de reciclar urea sanguínea hacia el rumen. Esta urea llega principalmente por vía salivar y en menor medida por difusión a través del epitelio, aunque existen discrepancias. Algunos investigadores consideran que la prevalencia de cada vía es función del contenido en amoníaco del rumen, de forma que la entrada de urea vía salivar sería un mecanismo pasivo que depende principalmente de la concentración de urea plasmática y de la producción de saliva y por tanto del tipo de ración consumida. Sin embargo, la difusión a través del epitelio estaría controlada por la actividad ureolítica de las bacterias adheridas a la pared ruminal, esta actividad estaría regulada directamente por la concentración de amoníaco ruminal. El amoníaco liberado por hidrólisis de la urea puede ser utilizado normalmente por las bacterias ruminales. En vacas que reciban raciones bien diseñadas, la cantidad de nitrógeno reciclado equivale prácticamente al nitrógeno absorbido como amoníaco, con un balance próximo a cero. El reciclado adquiere mayor importancia en animales que reciben raciones no sincronizadas en energía fermentescible y nitrógeno degradable. En este caso, la urea de la sangre permite compensar momentáneamente el déficit ruminal de nitrógeno debido a su rápida disponibilidad.b) Extracción intestinal: la flora microbiana de los ciegos y colon utiliza para su crecimiento la urea sanguínea como principal fuente de nitrógeno (más del 50% del total utilizado), otras fuentes de nitrógeno son los restos bacterianos ruminales no digeridos y las secreciones endógenas y las células epiteliales de descamación. La urea entra al intestino grueso por difusión pasiva a través del epitelio, es hidrolizada a amoníaco y se incorpora a la proteína microbiana. La extracción es tanto mayor cuanto más residuos alimenticios fermentables lleguen a intestino grueso. Esta vía es importante como pérdida de urea de la sangre, ya que sólo es reabsorbido como amoníaco el 8% del nitrógeno extraído. Como ya dijimos, a este nivel no ocurre absorción de otros compuestos nitrogenados.c) Eliminación renal: más del 60% de la urea plasmática es eliminada vía renal, y esta cantidad representa aproximadamente el 85% del nitrógeno urinario. La eliminación renal es también la vía principal de eliminación de los compuestos nitrogenados no ureicos. Aproximadamente el 98% de la alantoína presente en orina deriva del catabolismo de las purinas microbianas. La creatinina y la metil-histidina se encuentran en una concentración media de 6 y 0.7 mg por kg de peso vivo, respectivamente.
Los compuestos nitrogenados eliminados en los productos de excreción tienen una utilidad marcada como indicadores de diversos procesos metabólicos. Así, por su facilidad de difusión en los líquidos orgánicos, la urea aparece en la leche y su medida puede ser utilizada para conocer la relación --energía fermentescible/proteína degradable-- de la ración que reciben el ganado. Curiosamente la urea presente en leche se relaciona estadísticamente con la urea procedente del exceso de proteína degradable pero no con la urea de origen metabólico endógeno. Por otro lado, la alantoína es un indicador fiable de los ácidos nucleicos microbianos que llegan a intestino delgado y en consecuencia su medida en orina o en leche sirve para la estimación directa de la proteína microbiana digerida, debido a la relación constante purinas/proteína. En el caso de la creatinina y la metil-histidina presentes en la orina el nivel basal representa el equilibrio en la renovación continua de la masa muscular del animal y por tanto su valoración permite estimar cambios en la tasa de degradación de la proteína muscular debidos a variaciones en el estado nutritivo.
La síntesis de urea (y la excreción) supone un coste energético al organismo. Estoquiométricamente se calcula un consumo de 4 moles de ATP por mol de urea producida aunque posiblemente sea una sobreestimación ya que en el proceso se produce fumarato que es un intermediario del ciclo de Krebs. En cualquier caso, la necesidad de eliminar grandes cantidades de amoníaco supone un gasto extra de energía al organismo y puede tener una repercusión importante en el balance energético, sobre todo en situaciones de elevada demanda de energía como ocurre al comienzo de la lactación. Por otra parte, si ocurre función hepática disminuida (lipidosis hepática), la ureagénesis se ve inhibida. En estas circunstancias, el amoníaco en exceso es detoxificado alternativamente mediante la síntesis de glutamina desde ácido glutámico. La síntesis de ácido glutámico consume a su vez alfa-oxoglutarato que es un precursor glucogénico intermediario del ciclo de Krebs. El alfa-oxoglutarato es desplazado muy eficazmente del ciclo de Krebs ante una presencia excesiva de amoníaco.
Por tanto, la detoxificación del amoníaco puede repercutir doblemente de forma negativa en el metabolismo energético: consumo de ATP y menor disponibilidad de precursores glucogénicos. La capacidad de detoxificación orgánica del amoníaco es superada cuando su concentración ruminal es mayor de 80 mg/100 ml, entonces puede ocurrir intoxicación. El cuadro clínico es debido a la alteración del equilibrio ácido-básico y a la alteración del metabolismo energético celular por agotamiento del alfa-oxoglutarato, lo que es particularmente manifiesto en las células nerviosas. Otro efecto contraproducente del exceso de urea en sangre es el efecto negativo sobre el estado de salud del animal y sobre la reproducción.
NECESIDADES PROTEICAS PARA EL MANTENIMIENTO.
Las necesidades proteicas para mantenimiento están compuestas por las pérdidas a través de la piel, las pérdidas ocurridas en la porción anterior del tracto digestivo, la pérdida obligada de nitrógeno en la orina y las pérdidas de proteínas endógenas en las heces.
Las pérdidas de nitrógeno dérmico incluyen la renovación de la capa (crecimiento de pelos y descamación epitelial) y otras pérdidas queratinosas, expresándose normalmente en función del peso vivo metabólico (peso vivo elevado a la potencia 0.75)
Las pérdidas de proteína endógena que acontecen en la porción anterior del tracto digestivo corresponden a mucoproteínas de la saliva, células epiteliales de tracto respiratorio, restos celulares procedentes de la abrasión del epitelio de la boca, esófago y retículo-rumen, restos similares de omaso y abomaso, y secreciones de abomaso. Probablemente, las tres primeras fracciones son degradadas por los microorganismos ruminales y no llegan a duodeno. La contribución de las restantes fracciones al conjunto de proteína presente en duodeno es importante y se expresa de forma proporcional al consumo de materia seca. Esta fracción proteica es una pérdida parcialmente compensada porque es redigerida en intestino delgado y se incorpora al conjunto de proteína metabolizable.
Las pérdidas de proteína endógena que acontecen en la porción anterior del tracto digestivo corresponden a mucoproteínas de la saliva, células epiteliales de tracto respiratorio, restos celulares procedentes de la abrasión del epitelio de la boca, esófago y retículo-rumen, restos similares de omaso y abomaso, y secreciones de abomaso. Probablemente, las tres primeras fracciones son degradadas por los microorganismos ruminales y no llegan a duodeno. La contribución de las restantes fracciones al conjunto de proteína presente en duodeno es importante y se expresa de forma proporcional al consumo de materia seca. Esta fracción proteica es una pérdida parcialmente compensada porque es redigerida en intestino delgado y se incorpora al conjunto de proteína metabolizable.
La pérdida de nitrógeno en la orina representa la diferencia entre la síntesis y la degradación de la proteína corporal. El continuo proceso de renovación de las proteínas es muy intenso (2-4% de la proteína corporal al día) de forma que, a cada instante, una parte de las mismas es degradada hasta aminoácidos y reemplazada por proteínas nuevamente sintetizadas. Aunque la masa proteica se mantiene, ocurre una pérdida inevitable de nitrógeno debida al catabolismo de parte de los aminoácidos liberados. Esta pérdida se denomina nitrógeno endógeno basal. La velocidad de renovación es función de la edad (mayor en animales más jóvenes), del órgano considerado (más elevada en tracto gastrointestinal e hígado, seguidos por los riñones, la piel y la musculatura), y por el tipo de proteína (mayor para las proteínas funcionales). Las pérdidas de nitrógeno endógeno basal se expresan comúnmente en función del peso vivo metabólico. Indicar que la pérdida de energía asociada a la renovación proteica supone aproximadamente un 15% de las necesidades energéticas de mantenimiento.
Las pérdidas fecales de proteína endógena se consideran debidas a la descamación de la mucosa posterior a intestino delgado y a las secreciones digestivas no reabsorbidas. La estimación de estas pérdidas es compleja debido a que las heces contienen otras fracciones nitrogenadas como restos alimenticios no digeridos, residuos microbianos indigestibles y células microbianas intactas. Estas pérdidas se expresan en función del consumo de materia seca.
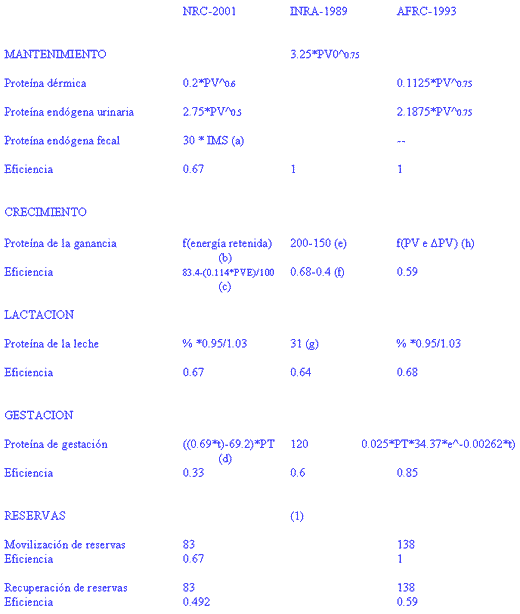
Debido a la dificultad práctica de mantener a los rumiantes en raciones libres de nitrógeno (reciclaje de urea plasmática a rumen y consumo disminuido de alimentos) y a la confusión entre las pérdidas urinarias y fecales de nitrógeno, la estimación de las necesidades proteicas de mantenimiento plantea amplias diferencias entre los diferentes grupos de investigación (CUADRO I). Para estimarla se siguen dos procedimientos:
CUADRO I / COMPARACION DE LA ESTIMACION DE NECESIDADES PROTEICAS ENTRE SISTEMAS.
Notas:
CUADRO I / COMPARACION DE LA ESTIMACION DE NECESIDADES PROTEICAS ENTRE SISTEMAS.
Notas:
Los valores son de necesidades netas en gramos por kilo
Para convertir las necesidades netas en proteína metabolizable hay que dividir por la eficiencia correspondiente, que está expresada en tanto por uno. PV : PESO VIVO (kg)
IMS: INGESTA DE MATERIA SECA (kg)
PVE : PESO VIVO EQUIVALENTE = RELACION ENTRE EL PESO VIVO ACTUAL Y EL PESO VIVO DE LA RAZA MODELO, CUYO PESO VIVO ADULTO ES 478 KG, LUEGO PVE= PESO ACTUAL *(478/PESO VIVO ADULTO)
DPV: INCREMENTO DE PESO VIVO (kg)
PT : PESO DEL TERNERO (kg)
t : DIAS DE GESTACION(a) valor en proteína metabolizable, es corregido por la fracción fecal correspondiente a la proteína microbiana = (30 * IMS) - (0.5*((PROTEINA METABOLIZABLE MICROBIANA/0.8)- PROTEINA METABOLIZABLE MICROBIANA). La proteína endógena que llega a duodeno supone un aporte de proteína metabolizable (=4.76 gramos * IMS) que debe recuperarse y equivaldría a = (8.9 gramos * IMS), por tanto el balance neto de proteína metabolizable endógena intestinal es de = -4.14 gramos * IMS
(b) La retención de proteína en el crecimiento es función de la energía retenida por kilo de peso ganado, y dicha energía es función de la edad y de la velocidad de crecimiento.
(c) La eficiencia es función de la edad, cuando se alcanza el peso vivo adulto es constante = 0.29
(d) Válido a partir de los 190 días de gestación.
(e) Movilización de reservas: de 5 a 7 kilos de proteína metabolizable en 60 días, y deben recuperarse a lo largo de la lactación
(f) El contenido y la eficiencia disminuyen con la edad
(g) Leche estándar: 3.1%
(h) Disminuye al aumentar el peso vivo y la ganancia de peso
=0.8*((168.07-0.16869*PV+0.0001633*PV^2)*(1.12-0.1223*DPV))
--Extrapolación a raciones libres de proteína: da valores muy bajos de nitrógeno urinario y mayores de nitrógeno endógeno fecal, aproximadamente 15% y 85% respectivamente para vacas en producción normalmente alimentadas.
--Mantener a los animales en raciones libres de nitrógeno pero adecuadas en los demás nutrientes que deben aportarse puros por infusión intragástrica, para evitar el reciclaje ruminal y a la extracción en intestino grueso. Los valores de nitrógeno endógeno urinario representan la mayor pérdida de nitrógeno (90-95%) frente a las pérdidas fecales.
Curiosamente cuando los animales reciben raciones de mantenimiento, la suma de las pérdidas urinarias y fecales calculadas por ambos procedimientos es muy similar. Sin embargo, al suministrar raciones de producción (mayor consumo de materia seca), las necesidades de mantenimiento son mayores para aquellos sistemas de alimentación que valoran el nitrógeno metabólico fecal en función del consumo. Como esta forma de cálculo es la que parece ajustarse más a los resultados productivos observados, la pregunta que se plantea es: ¿el incremento de las necesidades nitrogenadas en animales en producción es debido al incremento lineal de las necesidades de mantenimiento o a un incremento de la ineficacia de utilización de la proteína para los procesos productivos al aumentar el nivel de alimentación? En cualquier caso, la inclusión de las pérdidas fecales en el cálculo de las necesidades de mantenimiento supone que la estimación de las pérdidas urinarias debe reducirse en alguna medida.
En la estimación de la eficacia con que se usa la proteína metabolizable para el mantenimiento de la masa corporal también existen discrepancias entre los diferentes sistemas de alimentación (CUADRO I).
NECESIDADES PROTEICAS PARA EL CRECIMIENTO.
Cuando un animal recibe una ración adecuada en energía, el porcentaje de proteína de la ganancia de peso disminuye y el contenido de grasa aumenta al aproximarse a la madurez. La madurez química se alcanza cuando la ganancia de peso contiene cantidades mínimas de proteína. En el caso del ganado vacuno de raza frisona esto ocurre en torno a los 5 años de edad. Por otro lado, cuanto mayor es la velocidad de crecimiento, más aumenta la fracción de la ganancia que se retiene en forma de grasa en detrimento de la deposición de proteína. Por tanto, la estimación de la retención de proteína durante el crecimiento se hace en función de la retención de energía, cuyo valor recoge ambos aspectos (peso vivo y velocidad de crecimiento).
Para ajustar las necesidades proteicas para el crecimiento debemos considerar dos períodos claramente diferenciados:
a) Desde la primera concepción hasta que se alcanza el peso vivo adulto: la subalimentación nitrogenada durante la primera gestación y a lo largo de la primera lactación reduce la producción de leche en las lactaciones sucesivas (crecimiento diario de 0.69 y 0.104 kg/día, respectivamente). Sin embargo, las necesidades nitrogenadas para el crecimiento son cuantitativamente poco importantes en vacas lactantes a partir de la segunda lactación (crecimiento diario menor de 0.06 kg/día).b) Desde el nacimiento a la primera concepción: las necesidades de proteína para novillas en crecimiento se calculan de forma similar a los animales de engorde con la salvedad de que el ritmo de crecimiento no es máximo sino el adecuado para evitar engrasamiento corporal (crecimiento diario de 0.87 kg/día). Los aportes de proteína y energía durante la recría de novillas deben ser tales que el crecimiento de la masa corporal libre de grasa sea óptimo sin incremento del índice de estado de carnes. Además, la retención de energía en exceso provoca engrasamiento del parénquima mamario lo que reduce su funcionalidad posterior. Para conseguir este objetivo parece ser que la relación proteína/energía de la ración es más importante que simplemente la energía suministrada. Dicha relación debe ser adecuada y normalmente constante a lo largo del crecimiento, es decir, si se desea aumentar el ritmo de crecimiento, debe incrementarse el aporte de proteína.
La eficacia con que se utiliza la proteína metabolizable para la deposición de proteína es probablemente mayor en animales jóvenes (CUADRO I).
NECESIDADES PROTEICAS PARA LA PRODUCCION DE LECHE.
La producción de proteína láctea es cuantitativamente el mayor gasto nitrogenado de la vaca lechera. La proteína láctea se compone de una fracción de proteína verdadera (aproximadamente 95%) y diversidad de compuestos nitrogenados no proteicos. La fracción proteica es principalmente caseína (más del 80%). El principal compuesto nitrogenado no proteico es urea.
La ubre extrae los aminoácidos de la sangre con elevada eficacia según sus necesidades. Los aminoácidos esenciales se pueden dividir en dos grandes grupos: aquellos en que la cantidad extraída es similar a su presencia en la proteína sintetizada (metionina, fenilalanina, histidina y triptófano), y los restantes aminoácidos esenciales en que la extracción es mayor que su secreción en la leche. En conjunto, la extracción de aminoácidos esenciales es mayor que su presencia en la proteína láctea y lo contrario es cierto para los aminoácidos no esenciales. Los aminoácidos de cadena ramificada (isoleucina, leucina y valina) constituyen el 50% de los aminoácidos esenciales presentes en la proteína láctea.
La síntesis de proteína láctea se estima que consume el 11% del ATP generado en la ubre, si a esto le sumamos la síntesis de proteína endógena de la glándula mamaria (equivalente a 2.5 veces la síntesis de proteína láctea), el consumo total de energía para la síntesis de proteína en la ubre supone un 37% del ATP disponible a este nivel. Estos valores indican la influencia de la energía disponible en la síntesis de proteína láctea. De hecho la respuesta en proteína producida es mayor al incremento de energía consumida que al de proteína. Esta influencia ha sido recogida con gran sentido por los investigadores holandeses para expresar la eficacia de utilización de la proteína metabolizable para la síntesis de proteína láctea (CUADRO I).
NECESIDADES PROTEICAS PARA LA GESTACIÓN.
Al progresar la gestación las necesidades proteicas son más importantes que las necesidades energéticas, de forma que las primeras aumentan en un factor de 2.5 frente a un factor 1.5 para las segundas. La retención de proteína en los productos de la concepción (feto, placenta, cotiledones, útero grávido y fluidos). En los 2/3 iniciales de la gestación, las necesidades proteicas son debidas al crecimiento de los tejidos anejos y el aumento de los fluidos uterinos, cuya composición en materia seca es mayoritariamente proteína. Estas necesidades aún en el momento de máxima deposición son muy pequeñas respecto a las necesidades totales del animal. El crecimiento fetal es cuantitativamente más importante en el último tercio de la gestación, donde su crecimiento es prácticamente exponencial, y representa la mayor fracción de nitrógeno retenida. La proteína retenida en la ubre representa un pequeño gasto que sólo es significativo al final de la gestación. El aumento del volumen plasmático debido a la gestación conlleva una retención adicional de nitrógeno mínima. Las cantidades totales de proteína depositada son aproximadamente: 0.6 kg en fluidos, 0.4 kg en placenta y cotiledones, 1 kg en útero, 7.5 kg en un feto de 40 kg al nacimiento y unos 0.6 kg en la ubre.
La peculiaridad del metabolismo fetal de los rumiantes estriba en la elevada demanda de aminoácidos para su oxidación, que equivale prácticamente a la cantidad utilizada para la síntesis de proteína. La importancia de esta oxidación es aún mayor en casos de subnutrición energética. Este fenómeno determina que la eficacia de utilización de la proteína para la gestación se considere muy baja en algunos sistemas (CUADRO I)
NECESIDADES DE AMINOÁCIDOS.
Cuando falta un aminoácido para continuar la secuencia de la proteína que está siendo sintetizada, la síntesis se detiene ya que dicha secuencia está determinada genéticamente para cada proteína y no puede ser alterada. En este caso los aminoácidos sobrantes son catabolizados, ocurriendo pues un aprovechamiento subóptimo de los mismos y aumentando el amoníaco que debe ser eliminado. Por el contrario, si el perfil de los aminoácidos disponibles es similar al requerido por el animal, la eficiencia de utilización para la síntesis proteica es máxima, todos los aminoácidos son igualmente limitantes para la producción y el exceso de nitrógeno se minimiza.
Asumiendo una proporción constante entre los aminoácidos necesarios, los requerimientos pueden calcularse cuando se conocen las necesidades de un sólo aminoácido. Normalmente el perfil de aminoácidos se expresa referido a la lisina ya que suele ser el primer aminoácido limitante de la producción y no es usado para la síntesis de otros compuestos nitrogenados. Parece ser que el perfil de aminoácidos no esenciales tiene poca repercusión sobre la síntesis proteica. Las pruebas experimentales sugieren que cuando los aportes de aminoácidos totales se aproximan a las necesidades, los requerimientos de aminoácidos no esenciales se cubren antes que los de los aminoácidos esenciales, y si se absorben individualmente en menor cantidad de la necesaria, pueden ser sintetizados en cuantía suficiente sin que se afecte la producción. No obstante, se considera conveniente que el perfil de aminoácidos esenciales se exprese en porcentaje de las necesidades totales de proteína. Por tanto, conociendo las necesidades totales de nitrógeno pueden estimarse las necesidades de aminoácidos esenciales.
Con los conceptos anteriormente expuestos podemos definir la “proteína ideal” como la proporción perfecta entre los aminoácidos esenciales individuales y de éstos con el nitrógeno total requerido para una producción óptima.
Normalmente, se considera que la lisina es el primer aminoácido limitante de la producción en raciones donde predomina la proteína de los cereales y sus productos mientras que la metionina lo es en raciones basadas en soja. Es más probable que la metionina sea colimitante en la mayoría de las ocasiones ya que dicha estimación, al estar realizada sobre la base del aporte de aminoácidos de la fracción no degradable de las proteínas de los alimentos, no tiene en cuenta la diversidad de funciones metabólicas de la metionina. La isoleucina, la leucina, la histidina y la arginina serían los siguientes aminoácidos más frecuentemente limitantes.
A pesar de que no existen evidencias de que los aminoácidos no esenciales como grupo lleguen a ser limitantes, aún no se ha determinado el papel que pueden desempeñar algunos de ellos en determinados momentos productivos. La prolina y la glutamina han recibido especial atención debido a tres hechos: se encuentran en la caseína en mayor proporción que la proteína microbiana, su captación por la ubre es menor que su secreción en la leche y ambos son sintetizados desde arginina. La prolina se ha relacionado con un incremento de la producción de proteína láctea en la segunda mitad de la lactación. La glutamina participa intensamente en el metabolismo energético visceral especialmente durante los períodos de subnutrición energética. Por otro lado, la fenilalanina y la metionina contribuyen en un 50% a cubrir las necesidades de tirosina y cistina respectivamente. Se deduce que el aporte de los aminoácidos no esenciales mencionados podría suponer una reducción de las necesidades de los aminoácidos esenciales que participan en su síntesis.
La estimación cuantitativa de las necesidades de aminoácidos se puede realizar por dos vías:
a) Método factorial: con este método es necesario definir las necesidades netas de proteína para mantenimiento y cada una de las producciones, la composición en aminoácidos de los productos y la eficiencia de absorción y de utilización de los aminoácidos absorbidos. El principal problema de este método es que los errores cometidos en las estimaciones, particularmente en las eficiencias, son acumulativos y pueden generar resultados muy alejados de la realidad.b) Medida dosis-respuesta: aquí las necesidades son estimadas a partir de infusiones graduales del aminoácido a estudiar, de forma que se identifican respuestas productivas a las diferentes cantidades aportadas. Por este método, se ha calculado que la proporción óptima de lisina y metionina en la proteína metabolizable es aproximadamente de 7.3% y 2.5% respectivamente, expresado sobre aminoácidos esenciales las proporciones respectivas son 15% y 5%.
RESERVAS PROTEICAS.
Las vacas lecheras disponen de una cierta cantidad de proteína de reserva para su utilización como fuente de aminoácidos en condiciones de subalimentación como la que ocurre la comienzo de la lactación. Estas reservas son cuantitativamente pequeñas en comparación con las reservas grasas y por tanto muy limitadas en el tiempo. Se calcula que la capacidad máxima de movilización de proteína corporal al comienzo de la lactación es de unos 15-20 kg y que dicha movilización se prolonga hasta unas cinco semanas posparto. Por comparación, la movilización de grasa puede ser mayor de 80 kg y prolongarse más de 70 días posparto.
Normalmente, si la movilización máxima de nitrógeno no supera los 30 gramos diarios, la producción de leche no se ve afectada; esto se traduce en unas pérdidas de 5 a 7 kg de proteína corporal. Se estima que un kilo de tejido corporal movilizado por una vaca en condición corporal media contiene 600 gramos de grasa y únicamente 83 gramos de proteína. La proteína movilizada procede principalmente del tejido muscular y se estima que se utiliza con una eficacia del 80% como fuente de aminoácidos para la síntesis de proteína láctea.
Si la subalimentación nitrogenada persiste al avanzar la lactación ocurre disminución de la producción láctea y deplección de las reservas totales de proteína en el organismo con consecuencias en el estado de salud del animal. Este hecho es aún más grave si ha ocurrido déficit de proteína o energía durante la fase final de la gestación precedente. La repleción de las reservas proteicas ocurre con una eficiencia similar a la del crecimiento.
Por otro lado, el conjunto de urea plasmática sirve como fuente de nitrógeno rápidamente disponible para los microorganismos ruminales ante situaciones de deficiencia de nitrógeno degradable frente a la disponibilidad de energía fermentescible. El valor de este reciclado es tal que ante el suministro de raciones libres de nitrógeno, la fermentación ruminal se mantiene a ritmo decreciente y sólo cesa completamente en el curso de varios días. La cantidad de nitrógeno reciclado se ha calculado en función del porcentaje de proteína ingerida, mediante la regresión de los valores para diferentes consumos de proteína y materia orgánica digestible. En cualquier caso, en condiciones normales de alimentación la importancia del reciclado ruminal de nitrógeno es mínima ya que el balance neto (entrada - salida de amoníaco) es prácticamente cero.
CONCLUSION.
Las necesidades de proteína de los animales se estiman de forma factorial incluyendo las necesidades para mantenimiento, crecimiento, lactación, gestación y recuperación de las reservas proteicas. Las necesidades de proteína disponible para su absorción intestinal son una función de la proteína retenida en cada uno de los procesos mencionados mayorada por los gastos inherentes al proceso de síntesis. Dichos gastos constituyen la ineficacia. Para conseguir la máxima eficiencia de utilización de la proteína absorbida es necesario que su perfil de aminoácidos y la relación entre los aminoácidos esenciales y no esenciales sea óptima en función de la producción deseada. Por tanto, se hace necesario valorar también las necesidades nitrogenadas de los rumiantes como aminoácidos metabolizables, indicando las relaciones más adecuadas entre ellos y de los esenciales frente a los no esenciales.
Si la estimación de las necesidades nitrogenadas es estrictamente ajustada dentro de los conocimientos actuales es posible reducir la proteína total aportada en las raciones lo que reduce el riesgo sanitario, evita incidencias sobre la reproducción, aumenta la eficacia de la producción y reduce los costes de alimentación. Como factor de repercusión social hay que hacer hincapié en la posibilidad de reducir el nitrógeno presente en las excretas disminuyendo el impacto ambiental de este tipo de explotación.
Artículo publicado completo en Mundo Ganadero nº 145. Junio de 2002.© Mundo Ganadero. El mensual Mundo Ganadero lo edita Eumedia, S.A. en Madrid. C/ Claudio Coello, 16. 28001 MADRID.
Temas relacionados:
Autores:

Universidad de Córdoba - España
Recomendar
Comentar
Compartir

27 de octubre de 2008
Estimado Señor Martinez:
He leido con atención su completo artículo y he encontrado muy buena la explicacion en términos conceptuales lo referente al ciclo del nitrógenos, especialmente en el rumen.
Sin embargo quisiera solicitarle si pudiera establecer la necesaria relación requerida para efectos de producción microbiana, entre Proteina Degradable - o Nitrogeno degradable en rumen y los necesarios carbohidratos degradables en rumen.
Ojalá en terminos cuantitativos que es como realmente podemos aprovechar la información.
Atentamente y saludos.
Recomendar
Responder

Universidad de Córdoba - España
8 de marzo de 2007
Estimado Jorge:
Ha entrado hace poco en el comercio en España un producto a base de Nitrógeno No Proteico tratado químicamente para conseguir una lenta degradación ruminal. Mi opinión sobre este tipo de productos se fundamenta en dos cuestiones técnicas:
1ª) para que el rumiante pueda aprovechar eficientemente el Nitrógeno No Proteico, este debe ser degradado en el rumen, por tanto, ¿está equilibrada la velocidad de degradación de ese producto con la tasa de vaciado ruminal, o por el contrario existe el riesgo de que una gran parte del mismo pase a intestino donde ocasionará más problemas que beneficios?
2ª) el animal rumiante está adaptado a conservar nitrógeno para mantener la actividad microbiana ruminal, y dicho nitrógeno, cuando la ración es deficiente en el mismo, llega a los microorganismos en forma de urea salivar y transepitelial, procedente a su vez de la ineficiente utilización de la proteína de la ración, reduciendo además la excreción de nitrógeno en heces y orina; por tanto, ¿es necesaria la incorporación de Nitrógeno No Proteico, si las diferentes materias primas de la ración aportan suficiente cantidad de proteína bruta en general, y de proteína degradable en particular?
En resumen, me preocupa la relación entre la velocidad de degradación y la tasa de vaciado ruminal, ya que un desequilibrio entre ambas puede ser causar problemas, además de la ineficiente utilización del producto. Por otro lado, la utilización de fuentes de Nitrógeno No Proteico solamente está justificada en el caso de raciones extremadamente pobres en dicha fracción proteica, en cuyo caso, por otra parte, son un excelente complemento, siempre que se acoplen en la medida de lo posible a la energía fermentescible aportada por la ración.
Saludos,
Profesor Asociado del Dpto. de Producción Animal. Universidad de Córdoba (España)
Director Técnico de Fábricas de Pienso y Asesor de Explotaciones Ganaderas
Recomendar
Responder

Universidad de Córdoba - España
23 de febrero de 2007
Estimado Carlos Ernesto:
En próximos artículos haremos referencia a la forma más conveniente de suplir las necesidades proteicas del ganado vacuno lechero en relación a la doble vía de los rumiantes, esto es satisfacer las necesidades de los microorganismos ruminales con fuentes de proteína degradable que estén equilibradas con la energía fermentescible de la ración, estimulando de esta forma la producción de proteína microbiana, y por otro lado equilibrar dicha proteína con fuentes de proteína no degradable pero digestibles en intestino delgado para conseguir en definitiva el aporte de aminoácidos más adecuado (proteína ideal). Esta la forma más efectiva conocida hasta hoy de prevenir las deficiencias y los excesos de nitrógeno alimentario, sin perjudicar la salud de las vacas y favoreciendo su producción. Espero que sea de su interés.
Atentamente,
Andrés L. Martínez Marín
Profesor Asociado del Dpto. de Producción Animal. Universidad de Córdoba (España)
Director técnico de Fábricas de Pienso y Asesor de Explotaciones Ganaderas
Recomendar
Responder

10 de marzo de 2010
De verdad estoy muy agradecida con ustedes ke m hanayudado mucho en mi investigación que Dios los bendiga
Recomendar
Responder

22 de septiembre de 2009
Actualmente se me ha presentado un problema de sobreutilización de proteina en la dieta de vacas lecheras. Mi pregunta es: Puede producirse un sobreacondicionamiento corporal por el exceso de proteina, lo que se puede ver reflejado con baja de producción láctea??
Gracias
Recomendar
Responder

17 de septiembre de 2009
¿Quisiera saber las repercusiones del sobre acondicionamiento del ganado lechero en la reproducción?
Recomendar
Responder

8 de marzo de 2009
buenos dias
mi nombre es David, soy de Panama y quisiera que alguien me indique como puedo adquirir como especie de una tabla de las necesidaes diarias de los diferentes minerales de una vaca lechera en produccion e igualmente algo similar en cuanto a como poder seleccionar un buen producto en el mercado que contenga valores adecuados de los elementos reuqeridos, por ejemplo un producto que sea a base de calcio que cantidad minima debe decir en su etiqueta, y asi mismo los demas, fosforo, manganeso, magnecio, cobalto, selenio,etc, etc.me parece un poco dificil, pero si alguien me puede ayudar se lo agrdeceria muchisimo.
gracias,
David
Recomendar
Responder

13 de octubre de 2008
Me parece un excelente artículo en términos bioquímicos y metabólicos, muy orientado hacia la academia. Para nosotros los empresarios ganaderos es necesario que este tipo de artículos valiosos trasciendan a conclusiones prácticas, en este caso en cuanto a la nutrición proteica en las diferentes etapas del bovino, además de ilustrar la manera más adecuada y rentable de lograr una buena dieta asegurando la inclusión de aminoácidos esenciales.
Recomendar
Responder
6 de octubre de 2008
saludos amigos..
El ganado de leche también necesita de una sierta nutrición,necesita de proteínas y energía.
Entre algunas de las necesidades están la buena alimentacion,buena cantidad de proteina cruda,energía,grasa,lípidos,carbohidratos...esto es esencial para la buena nutricon y sobre todo salud de las vacas.
Recomendar
Responder

2 de octubre de 2008
Me pareció excelente la información vertida en su artículo, sin embargo, es estrictamente manejada con términos técnicos su redacción, por lo que sugiero que para los que no somos eruditos en la materia, se nos pudiera explicar en forma más detallada, qué elementos debemos utilizar para obtener los resultados mencionados. Quisiera que en una forma más sencilla nos iluminara con su sabiduría y pudiéramos entender más fácilmente cada uno de los pasos mencionados en su artículo.
Con información más detallada podríamos entender mejor lo explicado.
Agradezco su atención y espero abunde en el tema.
Gracias
Recomendar
Responder

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar
.jpg&w=3840&q=75)