Metabolismo de lipidos
Publicado: 1 de junio de 2018
Por: Mvz Humberto Troncoso Altamirano
Al menos en el hombre, entre el 95 y el 98 % del total de los ácidos grasos presentes en el plasma sanguíneo está contenido en los ésteres de ácidos grasos como los triglicéridos, los fosfolípidos y los ésteres del colesterol. Estos esteres de ácidos grasos se encuentran principalmente en forma de lipoproteínas plasmáticas. El resto, una pequeña porción de entre 2 y 5 %, se halla en forma no esterificada y está unido a un complejo albuminoide del plasma.
Las lipoproteínas realizan tres funciones principales: a) transportar las grasas de la dieta desde la mucosa intestinal, donde son absorbidas, hacia los tejidos del organismo animal; esta función la desempeñan los quilomicrones y los residuos de quilomicrones. b) transportar los triglicéridos desde el hígado hacia el resto de los tejidos del cuerpo, para almacenarse o ser oxidados para obtener energía. Las responsables de esta acción son las lipoproteínas de muy baja densidad (very low density lipoproteins), también conocidas como VLDL (por sus siglas en inglés). Una vez que las VLDL liberan los triglicéridos en los tejidos, los restantes constituyentes son devueltos al hígado en la forma de lipoproteínas de densidad intermedia (intermediate density lipoproteins), o IDL y también como lipoproteínas de baja densidad (low density lipoproteins), o LDL. c) actuar como mediador en el transporte inverso del colesterol; esta tarea recae en las lipoproteínas de alta densidad (high density lipoproteins), o HDL, y en las LDL, que devuelven al hígado el exceso de colesterol formado en los tejidos extrahepaticos. El cuadro 1, muestra la localización en donde tienen origen cada una de las lipoproteínas.
Los lípidos sanguíneos se transportan como lipoproteínas, que varían desde densidades muy bajas (VLDL), tales como quilomicrones hasta las de muy alta densidad (HDL). La densidad aumenta a medida que la proporción de proteínas en el complejo aumenta y a medida que los lípidos disminuyen. Los ácidos grasos libres (AGL), se transportan como un complejo con la albúmina.
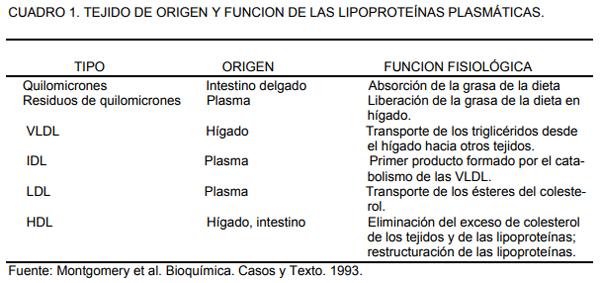
El cuadro 2, muestra la composición de las diferentes lipoproteínas plasmáticas en el hombre.
SINTESIS DE ACIDOS GRASOS.
El hígado, el tejido adiposo y la glándula mamaria son los tres sitios principales donde se lleva a cabo la biosíntesis de los ácidos grasos y los triglicéridos. El hígado es el órgano central para la interconversión y su metabolismo. La síntesis de ácidos grasos en el hígado, en el tejido adiposo y la glándula mamaria siguen vías parecidas; sin embargo, la actividad que cada una de ellas desarrolla varía de acuerdo con la especie animal. En las aves de corral (pollo para engorda y gallina de postura), el hígado es el órgano más activo; en el cerdo, el tejido adiposo muestra mayor actividad y, en los rumiantes, tanto el hígado, como el tejido adiposo y la glándula mamaria (en lactación) muestran la misma actividad.
Por otro lado, para los no rumiantes, el sustrato principal para la síntesis de ácidos grasos es la glucosa y, para los rumiantes (en general) el sustrato principal es el ácido acético proveniente de la actividad bacteriana en el rumen. En ambos casos, el exceso de estos sustratos son los que promueven a la síntesis de los ácidos grasos.
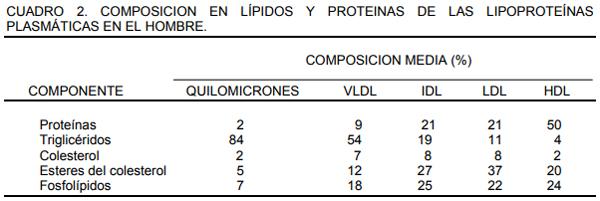
La síntesis de ácidos grasos se lleva a cabo en el citosol de las células activas y el producto activo para la síntesis es el acetil CoA proveniente de la glucosa vía glucólisis. A esta ruta también se le conoce como “síntesis de novo” o síntesis completa. El acetil CoA se sintetiza en el interior de las mitocondrias pero no puede salir hacia el citosol, por lo que se condensa con el oxalacetato que se difunde hacia el citosol.
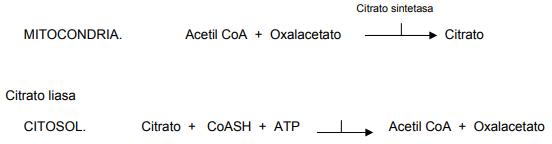
En el citosol, el acetil CoA tiene dos funciones importantes: es el iniciador de la síntesis de novo y, es la base para la elaboración de las unidades de malonil CoA. Malonil CoA es el donador de unidades carbonadas con las cuales crece el ácido graso en síntesis. Ambos productos entran en la ruta de la sintetasa de los ácidos grasos. La síntesis de un ácido graso se inicia por el extremo del carbono metileno (CH3) y termina en el extremo del carbono carboxílico (COOH).
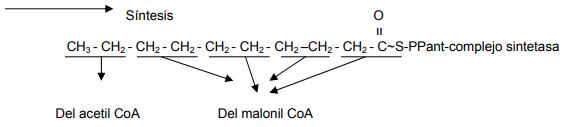
En la síntesis de ácidos grasos, el malonil CoA es sintetizado a partir de la carboxilación del acetil CoA. Esta reacción es mediada por un complejo enzimático, la acetil CoA carboxilasa, que contiene biotina. En esta reacción de carboxilación, actúa como intermediario el CO2 ligado covalentemente a la biotina unida a la enzima, formando un complejo llamado carboxibiotina. La reacción de carboxilación del acetil CoA a malonil CoA, es lo que regula la velocidad de la síntesis de los ácidos grasos.
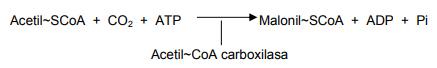
La síntesis de ácidos grasos tiene lugar en un complejo formado por siete enzimas independientes y una proteína transportadora que sujeta a la cadena acílica en crecimiento; a este complejo se le denomina ácido graso sintetasa. Es posible separar a este complejo enzimatico (de origen animal) en dos subunidades grandes aparentemente idénticas, las cuales están firmemente acopladas y actúan coordinadamente. Una subunidad contiene un grupo 4´-fosfopanteteína (que se conocerá posteriormente como PPant-SH, para fines didácticos), que proporciona un grupo sulfhidrilo al que se fija la cadena del ácido graso en crecimiento. El grupo 4´-fosfopanteteína es también el grupo funcional de la CoASH. La otra subunidad es el grupo cisteinil sulfhidrilo de la β-ceto sintetasa, conocida también como enzima condensadora (que se conocerá posteriormente como Cys-SH, para fines didácticos). Según este modelo, la cadena acil grasa (el ácido graso en crecimiento) va y viene entre el grupo PPant-SH y el grupo Cys-SH durante el proceso de síntesis.
Inicialmente, el grupo acetil del acetil CoA es transferido al grupo sulfhidrilo de PPant-SH. Esta reacción está catalizada por la acetil CoA-transacilasa:
A continuación, el grupo acetil, unido originalmente al grupo PPant, es transferido a un grupo cisteinil sulfhidrilo de la β-ceto sintetasa presente en la otra subunidad del complejo enzimático.
Así, se libera el grupo sulfhidrilo de PPant para aceptar al siguiente grupo que llega, un grupo malonil de la malonil CoA. La transferencia del malonil está catalizada por la malonil CoA aciltransferasa.
Enseguida, el residuo acetil es transferido de su sitio provisional en el grupo cisteinil sulfhidrilo de la segunda subunidad para que se condense con el residuo malonil ligado a PPant de la primera subunidad. En esta reacción, un grupo carboxilo se libera del malonato en forma de CO2, de manera que el producto resultante de esta condensación contiene cuatro átomos de carbono, en vez de cinco.
La condensación de los grupos acetil y malonil está catalizada por la β-ceto sintetasa. El proceso que impulsa dicha reacción es la descarboxilación del malonato. El producto de estas reacciones, el acetoacetil~S-PPant, es reducido a butiril~S-PPant por las restantes enzimas del complejo de la ácido graso sintetasa (la β-cetoacil reductasa, la enoil deshidratasa y la crotonil reductasa).
Tras esta serie de acontecimientos, se repite toda la secuencia, de tal manera que se van agregando pares de carbonos a la cadena arílica en crecimiento. En las reacciones anteriores, se formó el ácido butírico (un ácido graso de cuatro carbonos –C4-); la adición secuencial de pares de carbonos da lugar inmediatamente al ácido caproico (C6), después al ácido caprílico (C8), después al ácido cáprico (C10), y así sucesivamente hasta llegar a formar al ácido palmítico, un ácido graso de 16 carbonos.
La síntesis de un ácido graso por esta ruta llega a 16 carbonos todos saturados:
CH3-CH2-(CH2)12-CH2-COOH
Por lo anterior, la saturación de los carbonos de la cadena arílica se realiza tomando los hidrogeniones que aporta el NADPH que se genera por la vía de la pentosa fosfato, que es una de los principales generadores de este hidrógeno.
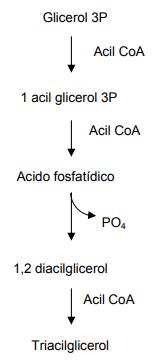
Los ácidos grasos generados por esta vía, no pueden vivir solos dentro del organismo, por lo que tienen que agruparse o condensarse. Para esto se lleva a cabo la síntesis de triacilglicéridos. Para esta síntesis se necesita de glicerol el cual proviene de la ruta de la glucólisis (a partir del glicerol 3P) al cual se le van adicionando por esterificación los ácidos grasos, como se observa a continuación:
Beta oxidación.
Los ácidos grasos almacenados como triglicéridos en el tejido adiposo son la principal fuente de energía para la mayoría de los animales estudiados. El catabolismo de los ácidos grasos se produce en el interior de las mitocondrias, mediante un proceso que se conoce como β-oxidación, en el que se van eliminando sucesivamente pares de carbonos (dos carbonos a la vez) del ácido graso. La eliminación es en forma de acetil CoA.
El catabolismo del ácido graso se inicia por el extremo carboxílico del mismo, mediante la eliminación de dos hidrógenos del carbono β (beta- C3 en la cadena arílica), formándose así un grupo cetónico. Por lo tanto, es el átomo de carbono β el que se oxida, de lo que se deriva el término de β-oxidación. Posteriormente, se produce una escisión (corte) entre los carbonos α y β y el fragmento de dos carbonos queda libre en forma de acetil CoA.
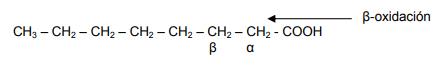
Una sola secuencia de β-oxidación que produzca un mol de acetil CoA, proporciona a la célula cinco moles de ATP cuando se oxidan en el ciclo de Krebs y, cada mol de acetil CoA proporciona a la célula 12 moles de ATP cuando se oxidan por la misma ruta. El siguiente es un ejemplo de la β-oxidación de un ácido graso de ocho carbonos (un ácido octanoico – C8:0, o ácido caprílico), como tioester de la CoA:
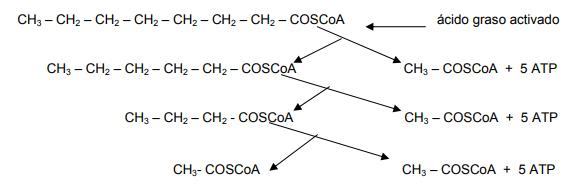
El primer paso en la ruta, es la activación del ácido graso, es decir, la formación de un tioéster de acil CoA por combinación con la CoASH. La enzima acil CoA sintetasa es la que regula la acción (también conocida como ligasa o tioquinasa).
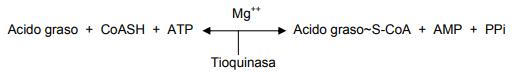
En particular, hay al menos cuatro acil CoA sintetasas independientes para los ácidos grasos que entran a β-oxidación: Una tioquinasa para ácidos grasos de cadena corta que activa al acetato y al propionato; una tioquinasa que activa ácidos grasos de cadena intermedia que contengan de 4 a 10 átomos de carbono; otra tioquinasa que activa a los ácidos grasos de cadena larga que contengan 12 ó más carbonos, y una tioquinasa específica para el ácido araquidónico (20:4ω-6). Las tioquinasas para los ácidos grasos de cadena corta e intermedia se encuentran en el interior de las mitocondrias, mientras que las enzimas que activan a los ácidos grasos de cadena larga y al ácido araquidónico se hallan en el retículo endoplásmico.
Para el caso de los ácidos grasos de cadena larga que van a degradarse por β- oxidación (como el ácido palmítico, que es el más común), una vez activado, no puede atravesar la membrana mitocondrial interna para llegar al lugar de la β- oxidación, por lo que para cruzarla el grupo acil de la CoASH se tiene que transesterificar a carnitina. Esta reacción es catalizada por la enzima carnitina aciltransferasa. Esta enzima existe en dos formas: a) la carnitina aciltransferasa I (CAT I), que se encuentra en la superficie externa de la membrana mitocondrial interna; b) una segunda forma de la enzima, la carnitina aciltransferasa II (CAT II), se encuentra en la superficie matricial de la membrana mitocondrial interna.
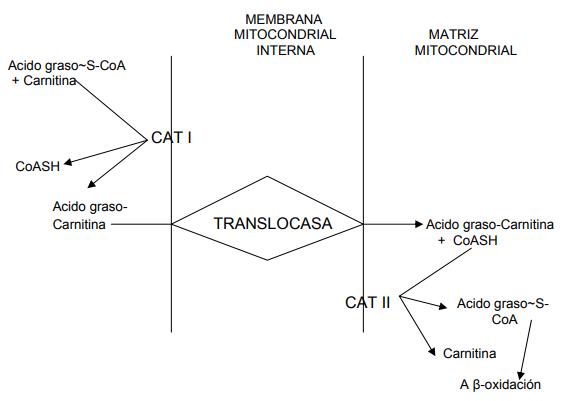
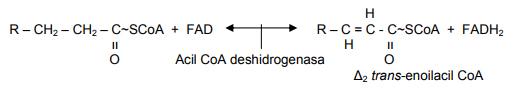
Una vez dentro de la mitocondria el ácido graso, se producen cuatro reacciones consecutivas: el primer paso es una deshidrogenación que requiere la presencia del dinucleótido de flavina adenina (FAD), y se forma un producto intermedio de acil CoA insaturado trans. FADH2 es transportado hacia la cadena respiratoria para su oxidación. La enzima que realiza esta reacción es la acil CoA deshidrogenasa.
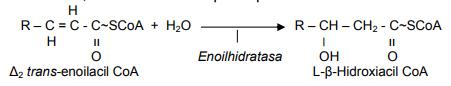
La segunda reacción es una hidratación del enlace insaturado, dando lugar a un L- β-hidroxiacil CoA, donde la enzima participante es la enoilhidratasa.
La tercera reacción es otra deshidrogenación que necesita de la presencia del dinucleótido de niacina adenina (NAD), que posteriormente es oxidado en la cadena respiratoria para su recuperación. La enzima que cataliza esta reacción es la L-β-Hidroxiacil deshidrogenasa:
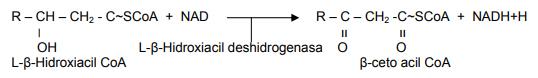
La cuarta y última reacción consiste en una escisión (corte) tiolítica y da lugar a un mol de acetil CoA y a un acil CoA que es un par de carbonos más corto que el ácido graso original que entró a β-oxidación.
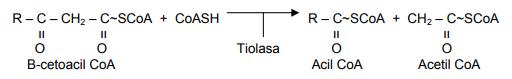
El acil CoA generado vuelve a entrar a β-oxidación y el proceso se repite hasta que toda la cadena es degradada a acetil CoA.
El FADH2 generado en el primer paso se oxida en la cadena respiratoria, produciendo 2 ATP, lo mismo que el NADH + H formado en el tercer paso se oxida igualmente en cadena respiratoria generando 3 ATP. Cada mol de acetil CoA generado por esta ruta se dirige hacia el ciclo de Krebs proporciona a la célula 12 ATP. En resumen, si un mol de ácido palmítico entrara a la ruta de la β-oxidación generaría:
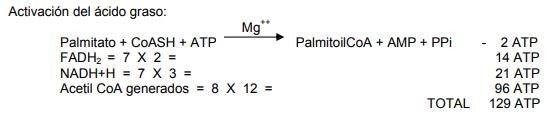
Para activar al palmitato se utilizan dos enlaces de fosfato de alta energía, uno en la reacción de la acil CoA ligasa y otro cuando se hidroliza el pirofosfato formado en esa reacción. En el ciclo de la β-oxidación el palmitoil CoA es convertido en ocho unidades de acetil CoA; para ello, se necesitan siete β-oxidaciones y cada secuencia de β-oxidación requiere de un FAD y NAD que generan cinco ATP.
LITERATURA CONSULTADA.
Church DC y Pond WG. Fundamentos de nutrición y alimentación de animales. Limusa-Noriega. México. 1990.
Maynard LA, Loosli JK, Hintz HF y Warner RG. Nutrición animal. 4a Ed. McGrawHill. México. 1981.
Montgomery R, Conway T y Spector A. Bioquimica. Casos y texto. 5ª Ed. MosbyYear book. España. 1993.
Huang WY, Kummerow FA. Cholesterol and fatty acid synthesis in swine. Lipids. 11(1):34-41. 1976.
Hansen HO, Knudsen J. Effect of exogenous long-chain fatty acids on individual fatty acid synthesis by dispersed ruminant mammary gland cells. J. Dairy Sci. 70(7):1350-1354. 1987.
Rahim A. Type of fatty acids, lipoprotein secretion from liver and fatty liver syndrome in laying hens. International J. Poultry Sci. 4(11):917-919. (2005).
Pearce J. Fatty acid synthesis in liver and adipose tissue. Proc. Nut. Soc. 42(2):263-271. (1983).
Temas relacionados
Autores:

Referentes que Recomendaron :
M.C. Fernando R. Feuchter A.Únete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicación
CADIA - Centro Argentino de Ingenieros Agrónomos
12 de junio de 2018
Muy completo y esclarecedor.
¿Que opina de la presencia de ácidos grasos insaturados, peroxidados, en las dietas de animales y humanos?¿que sucede con las enzimas?
Desde mya muchas gracias.
5 de julio de 2018
Excelente información

12 de junio de 2018
Felicidades una información muy clara y completa.



Te puede interesar
.jpg&w=3840&q=75)
Kemin Industries, Inc. - México