Características perinatológicas de la capacidad termorreguladora en ganado Holstein neonato
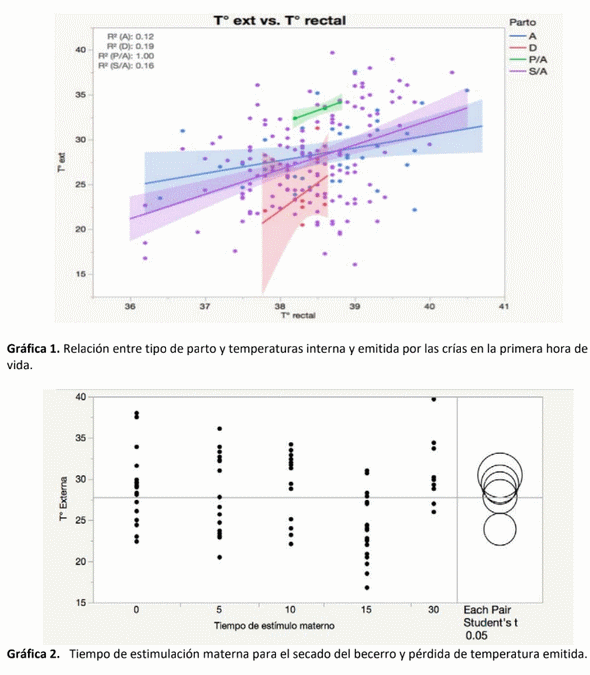
La atención perinatológica del recién nacido es un área deficiente en explotaciones lecheras en general. Se estima que del 30 al 50% de los becerros al parto requieren ayuda (House, 2015). El 75% de las muertes ocurren durante la primera hora de vida, estimándose que el 90% son previsibles. El impacto de una muerte neonatal es: económica (400-600 USD) (De Vries, 2006), genética, y de la siguiente generación, además de daño ginecológico en la madre (Mee, 2008). Se realizó un estudio observacional en el bajío mexicano, en una explotación lechera de Holstein- Friesian. El objetivo fue monitorear el sistema de termorregulación del recién nacido. El grupo cohorte fue de 43 becerros (17 machos y 26 hembras), los cuales recibieron atención durante el parto hasta la primera hora de vida. Se monitoreó la temperatura toraco-abdominal exterior, auricular y ambiental con una cámara termográfica de rayos infrarojos, y rectal con termómetro digital. Los resultados del análisis mostraron que los machos presentan una mayor pérdida de temperatura, en relación directa con mayor peso al nacimiento. Las menores pérdidas de calor fueron cuando el becerro fue estimulado por 15 minutos mediante el secado de pelaje por su madre. Inmediatamente después del parto, se identificó una caída de temperatura rectal de 0.6°C en promedio y externa de 3°C, a los 15 minutos la temperatura aumentó, siendo estadísticamente diferente al T°0 y <30 minutos después (Valor p <0.0001). Esta pérdida de temperatura corporal indicó que, pese a temperaturas ambientales de 34°C, existe la necesidad de estimular la circulación del becerro, proveerle una fuente de calor y abrigo, y alimentarlo prontamente con calostro para incrementar su energía y evitar hipotermia.
se evapora y el calor que se disipa desde las vías respiratorias, que al ser una actividad endotérmica le resta calor al becerro. La segunda ruta es la no evaporativa; ésta se lleva a cabo por medio de la transferencia de calor por conducción, es decir, el calor que pasa a través de la piel, lo que involucra el área en donde se encuentra, el tipo de cama y el ambiente (Cook et al. 2004). Ambas rutas de pérdida de calor involucran el uso de calorías, provocando así que el animal utilice las reservas con las que nace y con la consecuente hipotermia.
producción de lactato-L, como consecuencia de la saturación de CO2 que se mantiene por 4 horas en niveles elevados. La acidosis inicialmente respiratoria y se hace metabólica, cuando el lactato-L circulante no es metabolizado rápidamente por el riñón (Bleul y Gotz, 2013; Romano y Vázquez-Flores, 2018)). Una hipoxia prolongada, aunada con acidosis respiratoria y metabólica, genera somnolencia, el becerro se postra, disminuyéndose las probabilidades de sobrevivencia y reduciendo la absorción de inmunoglobulinas (Besser y Gay, 1994).
pélvicos y la relajación del cérvix. La hembra al parto, se manifiesta inquieta, puede presentar pérdida de apetito, y voltea la cabeza constantemente hacia el vientre cuando hay una contracción.
vez que el cérvix está completamente dilatado, la cría es expulsada en 22 minutos. Actualmente, con la condición genética de producir una cría chica al parto, por medio de toros que tienen la característica de facilidad de parto, los tiempos se han disminuido considerablemente de 20 a 40 minutos.
becerro como el peso, duración de la gestación, sexo, raza, nutrición y clima. Y los propios de la madre como el tamaño de la pelvis, peso corporal al servicio efectivo, edad de la hembra y condición corporal al parto (Mee, 2011).
infrarrojas en tiempo real (imagen 1). La pérdida de calor no evaporativa involucra el flujo de calor generado por el metabolismo, el organismo mantiene la homeostasis por efecto de la radiación, convección y conducción. Entendiendo la radiación como el balance neto entre la ganancia de calor por radiación solar y la pérdida de calor por contacto de la piel con la superficie del piso. La transferencia de calor por conducción es el intercambio de calor desde el pelaje hacia el ambiente. La pérdida de calor por convección, se refiere al paso del aire sobre el pelaje que cuando está húmedo permite el enfriamiento de la superficie por el intercambio de temperatura entre el aire y pelaje (Kasari, 1994). Es por esto que se realizaron tomas de temperatura por medio de la cámara termográfica, además de la temperatura rectal que evalúa la respuesta interna al ambiente (Vázquez-Flores y Lucio-Rodríguez, 2017).

haber transcurrido un máximo de 15 minutos con la madre, al becerro era retirado del área de parto y se transportado al cunero.
2. Poca Ayuda (P/A)
3. Ayuda (A)
4. Distocia (D)
análisis demográficos, medias y desviaciones estándar, pruebas paramétricas de T-de Student, Hsu ´s MCB, y regresión lineal.
- RESULTADOS
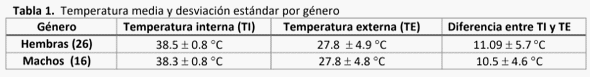
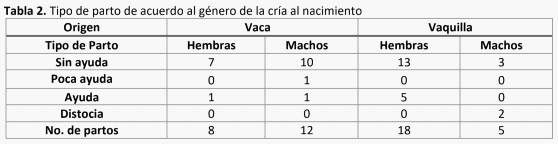
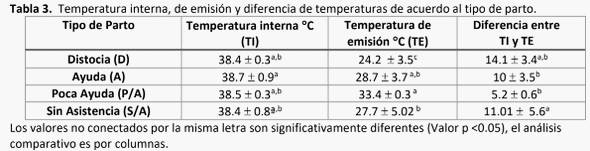
donde perdieron menos temperatura en general, no hubo diferencias en las hembras (Gráfica 2).
alimentarlo prontamente con calostro para incrementar su energía y evitar hipotermia.
1. Alexander, G., Bennett, J. W., Gemmell, R. T. (1975). Brown adipose tissue in the new-born calf (Bos taurus). The Journal of Physiology, 244(1), 223-234.
2. Asakura, H. (2004). Fetal and neonatal thermoregulation. Journal of Nippon Medical School, 71(6), 360-370.
3. Atrian, P., & Shahryar, H. A. (2012). Heat stress in dairy cows (a review). Research in Zoology, 2(4), 31-37.
4. Barrier, A., Mason, C., Dwyer, C., Haskell, M. & Macrae, A. (2013). Stillbirth in dairy calves is influenced independently by dystocia and body shape. The Veterinary Journal 197(2): 220-223.
5. Barrier, A. C., Ruelle, E., Haskell, M. J., & Dwyer, C. M. (2012). Effect of a difficult calving on the vigour of the calf, the onset of maternal behaviour, and some behavioural indicators of pain in the dam. Preventive Veterinary Medicine, 103(4), 248-256.
6. Beam, A. L., Lombard, J. E., Kopral, C. A., Garber, L. P., Winter, A. L., Hicks, J. A., Schlater, J. L. (2009). Prevalence of failure of passive transfer of immunity in newborn heifer calves and associated management practices on US dairy operations. Journal of Dairy Science 92(8): 3973-3980.
7. Bellomo, R. (2002). Bench-to-bedside review: lactate and the kidney. Critical Care, 6(4), 322.
8. Berglund, B., Steinbock, L., Elvander, M. (2003). Causes of Stillbirth and Time of Death in Swedish Holstein Calves Examined Post Mortem. Acta Veterinaria Scandinavica 44(3): 111-120.
9. Besser, T. E., & Gay, C. C. (1994). The importance of colostrum to the health of the neonatal calf. Veterinary Clinics of North America: Food Animal Practice, 10(1), 107-117.
10.Bleul, U., & Götz, E. (2013). The effect of lactic acidosis on the generation and compensation of mixed respiratory-metabolic acidosis in neonatal calves. The Veterinary Record, 172(20), 528-528.
11.Carstens, G. E. (1994). Cold thermoregulation in the newborn calf. Veterinary Clinics of North America: Food Animal Practice, 10(1), 69-106.
12.Colburn, D. J., Deutscher, G. H., Nielsen, M. K., and Adams, D. C. (1997). Effects of sire, dam traits, calf traits, and environment on dystocia and subsequent reproduction of two-year-old heifers. Journal of Animal Science, 75(6), 1452- 1460.
13.Cook N. B., Nordlund KV (2004). “Veterinary Clinics of North America: food animal practice. Managing the transition cow to optimize health and productivity” Elsevier Saunders editors.
14.Collier, R. J., G. E. Dahl, and M. J. VanBaale. 2006. Major advances associated with environmental effects on dairy cattle. J. Dairy Sci. 89:1244–1253. Dahl, G., Tao S, Monteiro A. (2016). Effects of late-gestation heat stress on immunity and performance of calves, Journal of Dairy Science, Vol. 99 No. 4.
15.FLIR tools, versión 2.1 (2014) http://www.flir.com.mx/instruments/display/?i d=54865
16.Grove-White, D. (2000). Resuscitation of the newborn calf. In Practice, 22(1), 17-23.
17.Johanson, J. M. Berger., P. J. (2003). Birth Weight as a Predictor of Calving Ease and Perinatal Mortality in Holstein Cattle. Journal of Dairy Science 86(11): 3745-3755.
18.Kasari, Thomas R., and Steven E. Wikse. (1994). "Perinatal mortality in beef herds." Food animal practice.
19.Kadzere, C. Murphy M, Silanikove N, Maltz E., (2011). Heat stress in lactating dairy cows: a review. Livestock Production Science 77: 59– 91Mee, J. F. (2008). Prevalence and risk factors for dystocia in dairy cattle: A review. The Veterinary Journal, 176(1), 93-101.
20.Hovinen, M., Siivonen, J., Taponen, S., Hänninen, L., Pastell, M., Aisla, A. M., & Pyörälä, S. (2008). Detection of clinical mastitis with the help of a thermal camera. Journal of Dairy Science, 91(12), 4592-4598.
21.House, J. (2015). The Peripartum Ruminant. In Large Animal Internal Medicine (Ed B. P. Smith). Cap 283. St. Louis, Missouri: Elsevier.
22.Martin S. W., Schwabe C. W. and Franti C. E. (1975). Dairy Calf Mortality Rate: The Association of Daily Meteorological Factors and Calf Mortality. Can J Comp Med. Oct; 39(4): 377–388.
23.Mee, J. F., Berry, D. P., & Cromie, A. R. (2008). Prevalence of, and risk factors associated with, perinatal calf mortality in pasture-based Holstein-Friesian cows. Animal, 2(4), 613-620.
24.Mee, J. (2011). Bovine Neonatal Survival Is Improvement Possible. WCDS Advances in Dairy Technology, 23, 161-174.
25.Mee, J. F. (2013). Why do so many calves die on modern dairy farms and what can we do about calf welfare in the future? Animals, 3(4), 1036- 1057.
26.Miedema, H. M., Cockram, M. S., Dwyer, C. M., & Macrae, A. I. (2011). Behavioural predictors of the start of normal and dystocic calving in dairy cows and heifers. Applied animal behaviour science, 132(1), 14-19.
27.McGuirk SM: Reducing dairy calf mortality. Proc Am Assoc Bov Pract 40:126-131, 2007.
28.Nagy, D. W. (2009). Resuscitation and critical care of neonatal calves. Veterinary Clinics: Food Animal Practice, 25(1), 1-11.
29.Phillips, P. K., & Heath, J. E. (2001). An infrared thermographic study of surface temperature in the euthermic woodchuck (Marmota monax). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 129(2-3), 557-562.
30.National Research Council. (2001). Nutrient requirements of dairy cattle: 2001. National Academies Press.
31.Romano Muñoz, JL., Vázquez-Flores, S. Haubi Segura, CU, Shimada Miyasaka, A. (2018). Capítulo 9. Desórdenes metabólicos del rumiante en el libro de Nutrición Animal. 4a Edición. Editor Armando Shimada Miyasaki. Publicado. Editorial Trillas SA de CV, México. pp 277-306. ISBN 978-607-17-31201.
32.Rogers, P. A. M. (1999). Emergency acupoint Renzhong (GV26): A bibliography and review from textbook sources. In Proceedings of the 25th Annual International Congress on Veterinary Acupuncture (pp. 83-89).
33.Van Amburgh, M, (2017). Dairy replacement heifers: Transitioning from weaning through the first lactation, ADSA Discover 32 Conferences.
34.Vasseur, E., Borderas, F., Cue, R. I., Lefebvre, D., Pellerin, D., Rushen, J., Wade, K.M. and De Passillé, A. M. (2010). A survey of dairy calf management practices in Canada that affect animal welfare. Journal of Dairy Science, 93(3), 1307-1316.
35.Vasseur, E., Pellerin, D., de Passille, A. M., Winckler, C., Lensink, B. J., Knierim, U., & Rushen, J. (2012) Assessing the welfare of dairy calves: outcome-based measures of calf health versus input-based measures of the use of risky management practices. Animal Welfare, 21(1), 77-86.
36.Vázquez-Flores, S. and C. Lucio-Rodriguez (2017) Early non-invasive clinical diagnosis of hoof ulcers by infrared thermographic images (IRT) in milking dairy cows. ISSN 0022-0302, Journal of Dairy Science, 100 (Supplement 2): 27
37.Vermorel, M., Dardillat, C., Vernet, J., & Demigne, C. (1983). Energy metabolism and thermoregulation in the newborn calf. In Annales de Recherches Veterinaires (Vol. 14, No. 4, pp. 382-389).
38.Vieira, A. D. P., Guesdon, V., De Passille, A. M., von Keyserlingk, M. A. G., & Weary, D. M. (2008). Behavioural indicators of hunger in dairy calves. Applied Animal Behaviour Science, 109(2), 180-189.





