La cantidad de materia orgánica microbial (MOM), asi como el nitrógeno microbial (NM) que fluyó a duodeno fue calculado en base a los análisis de la bacteria aislada en el fluído ruminal, asi como de las muestras de duodeno obtenidas, usando purinas como marcadores microbianos 10.La materia orgánica fermentada (MOF) en rumen fue calculada como el resultado obtenido de restar a la MO consumida la diferencia cuantitativa observada entre la cantidad total de MO menos la MOM que ingresó a duodeno [(MOF = MOC-(MO-MOM)].
El N consumido que escapó a la digestión ruminal fue considerado al equivalente del total de N que ingresa a duodeno menos la suma de las cantidades de N amoniacal y N microbial que fluyó a duodeno.
La producción de metano en rumen se calculó a partir del balance teórico de fermentación en base a la distribución molar de AGV observada en el fluído ruminal14 y de la desaparición de MO en rumen.
Resultados
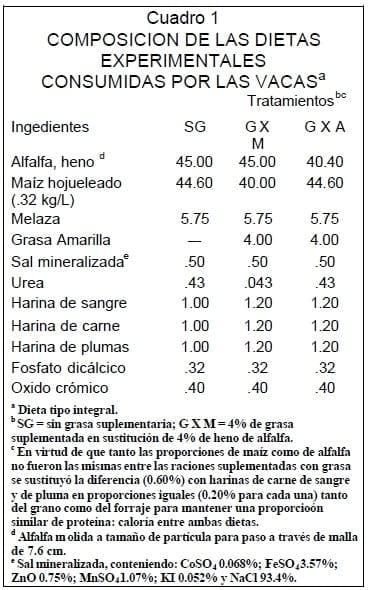
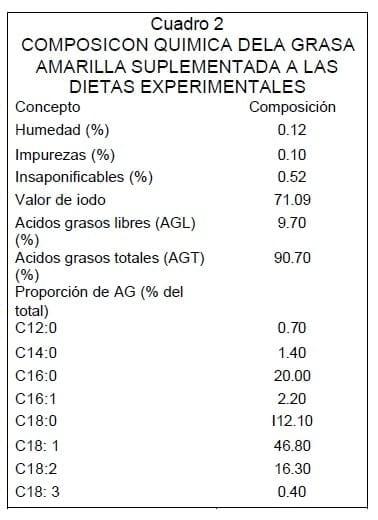
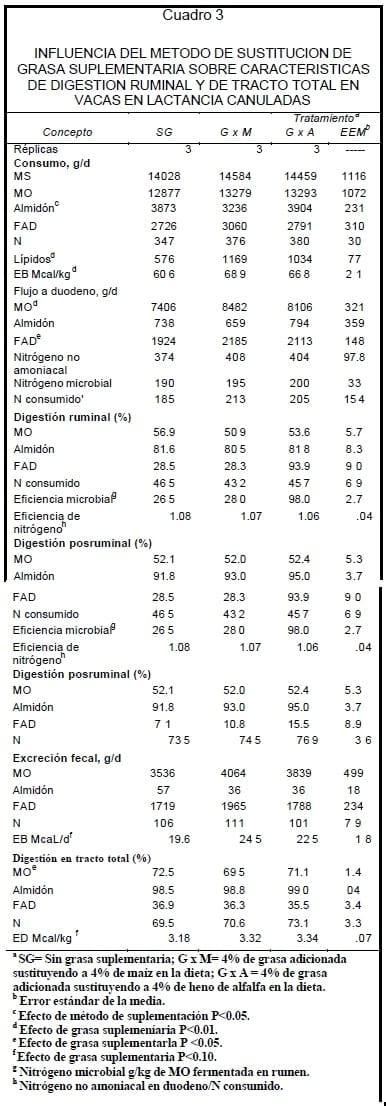
La característica de la grasa suplementada a las dietas experimentales se muestra en el Cuadro 2. Los valores de ácidos grasos libres (AGL), ácidos grasos totales (AGT), así como su perfil de ácidos grasos (AG) están dentro los rangos especificados para este tipo de grasa de acuerdo con las reglas y normas establecidas por la AFOA18. El efecto de los tratamientos sobre consumo de MS y características de digestión se muestra en el Cuadro 3. No hubo diferencia (P>0.20) en el consumo de MS dada por los tratamientos y debido a que no existió la misma proporción de forraje y maíz hojueleado entre los tratamientos 2 y 3 sucedió una diferencia en el consumo de almidón en 16.7% ( P<0.01) y de 8.8% (P>0.15) de FAD aunque este último no fue significativo estadísticamente.
En este sentido, la suplementación con grasa a los tratamientos
G x M y G x A reflejó una diferencia en el consumo de lípidos (1102 vs 576 g/d )de estas dietas en relación con el tratamiento SG, lo anterior representó un consumo 11.9% mayor (P<0.01) de EB (Mcal/d) para este tipo de dietas. No hubo efecto (P>0.10) de grasa suplementaria o método de suplementación, sobre digestión ruminal y posruminal de MO, N y almidón, aunque el método de suplementar grasa en sustitución de forraje tendió a disminuir en 18% (P> 0.20) la digestibilidad ruminal de FAD.
La suplementación con grasa favoreció (P<0.10) un mayor flujo a duodeno de N consumido en 13%, sin cambios en el flujo de nitrógeno no amoniacal (NNA) y NM. La eficiencia microbial (N microbial g/kg de MO fermentada), y eficiencia de N (NNA en duodeno/ N consumido) no fueron afectadas (P>0.20) por la adición de grasa a la dieta, o metodo de suplementación de la misma. El adicionar grasa a las dietas disminuyó (P<0.05) la digestibilidad de MO en tracto total en 3.0% , e incrementó (P<0.05) en 4.5% la energía digestible (ED,Mcal/kg) de la dieta.
El valor de la ED (Mcal/kg) para la grasa suplementada promedió 7.08, y resultó en 7.54 Mcal/kg cuando fue sustituida por maíz, y 6.62 Mcal/kg cuando se sustituyó por forraje, de acuerdo con el valor de la ED observado para la grasa en este experimento, la digestibilidad de la misma promedió 68%.
Los efectos de los tratamientos sobre pH ruminal, proporción molar de AGV y producción de métano en rumen se muestran en el Cuadro 4. El pH ruminal promedió 6.08 y no fue afectado (P>0.20) por los tratamientos. La adición de grasa a las dietas contribuyó a la disminución (P<0.10) en 13% a la proporción de propionato a nivel ruminal, sin afectar los otros parámetros evaluados a este nivel.
Discusión
El consumo en base MS (kg/d) no fue afectado por la suplementación de grasa o método de suplementación. Otras investigaciones indican que vacas suplementadas con grasa en niveles de 2.5% 19 hasta niveles igual o mayor al 5% 4,20,21,22,23 no tuvieron cambios en su consumo con respecto del testigo. En contraste , suplementaciones con 5% de grasa amarilla han denotado tanto disminución 24 como incremento en el consumo de MS 25. Muchos factores deben de considerarse para las inconsistencias mostradas en el consumo de MS cuando las dietas son suplementadas con grasas, esos factores han sido previamente discutidos por Coppock y Wilks26, asi como por Plascencia y Zinn27.
Contrario a nuestra suposición, el sustituir grasa por forraje en la dieta no se tradujo en efectos significativos sobre los parámetros aqui evaluados, aún cuando el consumo de almidón para
G x M fue en promedio diario 652 g menor con respecto a las dietas control y G x A. Lo anterior indica, que la cantidad de almidón removido en las dietas cuando se suplementa grasa en sustitución del concentrado dentro de los niveles recomendados (<5% de la ración) no es un factor determinante en la disminución de la capacidad de digestión de la fracción fibrosa de las dietas o de la síntesis microbial de N ,generalmente observado cuando se realiza esta práctica.
Los resultados sobre digestibilidad ruminal y de tracto total de MO y almidón obtenidos en el presente experimento concuerdan con estudios previos en donde grasas no protegidas han sido añadidas hasta en 5% a dietas altas en forrajes 5,28,29. Sin embargo, los efectos de la grasa suplementaria sobre la digestión ruminal de FAD han sido inconsistentes, ya que en algunos casos 1,29,30,31,32 adicionar grasa no tiene efecto negativo sobre la digestión de la fibra, aunque en otros estudios 3,33,34,35 la utilización de grasa disminuye en forma drástica su digestión ruminal. Zinn y Plascencia 36 indican que la base para esta inconsistencia aparente no está relacionada con la composición de la dieta en sí. Al parecer los efectos sobre la digestión de la fibra está más estrechamente relacionada con la proporción de ácidos grasos libres (AGL) contenidos en la grasa suplementada.
De cualquier forma, basándonos en éste, asi como en resultados de estudios previos30,31,33,36, se concluye que la grasa amarilla a diferencia de otras fuentes de grasas, tiene un efecto limitado sobre la digestión ruminal y de tracto total de la fibra.
El incremento (P<0.05) en 34 g/d de flujo de N dietético a duodeno, fue un reflejo directo de la diferencia en el consumo promedio diario de MS, al igual que de la cantidad de N ingerido (+31 g/d) para las dietas suplementadas con grasa; en consecuencia, este aumento no se reflejó en la digestibilidad ruminal del mismo. La mayoría de los estudios no indican efectos sobre la digestibilidad de N a nivel ruminal y de tracto total por efecto de la suplementación con lípidos a las dietas22,32,37,38,39. Aunque existen estudios donde se indica un efecto considerable de las dietas suplementadas con grasas en la digestión de N21,40.
El incremento en la digestibilidad de N en los estudios donde se suplementa grasa puede atribuirse principalmente a la práctica generalizada de incluir pasta de soya (proteína de alta degradabilidad ruminal)a las dietas con grasa para mantener la relación proteína:energía respecto de la dieta testigo. En el presente estudio, en las dietas suplementadas con grasa, las sustituciones se realizaron con fuentes de baja degradabilidad ruminal41,42(Cuadro 1) ,eso último explica el resultado.
En el presente experimento, así como en la mayoría de estudios previos, la suplementación de lípidos no tuvo efecto sobre el flujo a duodeno de NNA32,33,37,43,44. De igual forma, la ausencia de efectos dado por la suplementación de grasa sobre la eficiencia microbial y eficiencia de N han sido observados en otros estudios36,39,45, aunque varios trabajos30,46,47,48 indican aumentos en esos parámetros. En ese sentido, Doreau y Ferlay37 en un estudio recapitulativo, analizaron 38 comparaciones de 24 estudios donde se evaluó el efecto de la suplementación con lípidos sobre metabolismo de N en rumiantes, observando que el aumento en eficiencia microbial resulta sólo cuando existe una disminución de la digestión ruminal de MO por encima del 10% con respecto a las dietas no suplementadas.
Por otra parte, los aumentos en la eficiencia microbial observada en ocasiones con grasas suplementarias se considera como consecuencia en la disminución de la población de protozoarios ruminales49,50; sin embargo , la ausencia de cambios en el número de protozoarios en este tipo de dietas han sido indicados por Ohajuruka et al.21 y Towne et al.51
La suplementación de grasa incrementó (4.5%;P<0.10) la ED (Mcal/kg) de la dieta. Como consecuencia de que la ED del maíz hojueleado y del heno de alfalfa es de 3.88 y de 2.65 Mcal/kg, respectivamente (NRC52), el valor de la ED de la grasa suplementaria puede estimarse por medio de la técnica diferencial como sigue:
ED de la grasa prueba en GxM = [((ED de la dieta prueba - ED de la dieta control)/0.046 + 3.88 - (0.13 * 2.83)/0.87)].
ED de la grasa prueba en GxA = [((ED de la dieta prueba - ED de la dieta control)/0.046 + 2.65 - (0.13 * 2.83)/0.87)].
donde las constantes 3.88 y 2.65 son los valores de ED (Mcal/kg ) del maíz y heno de alfalfa, respectivamente (NRC52). Las constantes 0.13 y 0.87, corresponden a la proporción de las harinas de carne, pluma y sangre, así como de grasa suplementaria, en la cantidad total de grasa más las harinas proteínicas, las cuales remplazaron al maíz hojueleado o al heno de alfalfa en la dieta basal (Cuadro 1). De acuerdo con lo anterior, el valor de ED para grasa suplementaria fue de 7.54 y 6.62 Mcal/kg para G x M y G x A respectivamente, promediando 7.08 Mcal/kg. Considerando que la grasa contiene 9.5 Mcal/kg de EB, y la ED de la grasa fue de 7.08 Mcal/kg, entonces la digestibilidad aparente de la grasa resultó 74.5% a nivel de consumo ajustado a mantenimiento, o 68% a nivel del consumo mostrado en este experimento. En trabajos previos con dietas de finalización para novillos, se ha determinado un valor promedio de ED para grasa amarilla de 7.7 Mcal/kg30,53 ; por lo tanto, el valor obtenido aquí corresponde al 92% de ese valor. En una serie de estudios (20 observaciones en total) realizados por Zinn34, éste observó que la digestión posruminal de la grasa suplementaria (DL,%) en novillos está fuertemente asociada al consumo total de lípidos, expresando lo anterior con la siguiente ecuación: DL, % = 83.18 - 4.52LC - 0.68 LC3, donde LC = lípidos consumidos (g/kg PV). Aplicando lo anterior a los resultados de este trabajo, la digestión intestinal de lípidos esperada para las dietas suplementadas con grasa es 67%, valor muy cercano al 68% calculado en base a la ED observada para la grasa suplementaria usada en este experimento.
El resultado obtenido en los parámetros de fermentación ruminal con respecto a la disminución en un 13% de la proporción molar de propionato en las dietas suplementadas con grasa es incierto. En general la suplementación con grasa en dietas altas en forraje (>30%) no tiene influencia en el pH ruminal 54,55, o en la proporción molar de AGV 22,36,48,55,56,57,58. Aunque existen estudios con dietas similares donde la grasa aumenta la proporción de propionato y disminuye la proporción de acetato 59,60 no hay informes de que los lípidos suplementarios disminuyan la concentración molar del propionato ruminal.
Con base en lo anterior, se concluye que el suplementar con 4% de grasa a las raciones para ganado en lactación, puede efectuarse sustituyendo, en la misma proporción, al grano o al forraje sin efectos negativos sobre la fermentación ruminal, o sitio y tasa de digestión de MO, almidón, N, FAD y lípidos. Sin embargo, el valor nutrimental de la grasa suplementaria se redujo en 12% cuando ésta se adicionó sustituyendo al forraje.
Referencias
1. Jenkins TC. Nutrient digestion, ruminal fermentation, and plasma lipids in steers fed combination of hidrogenated fat and lecithin. J Dairy Sci 1990;73:2934-2939.
2. Devendra CA, Lewis D. The interaction between dietary lipids and fiber in the sheep. Anim Prod 1974;19:67-76.
3. Zinn RA, Plascencia A. Interaction of whole cottonseed and supplemental fat on digestive function in cattle. J Anim Sci 1993;71:11-17.
4. Jenkins TC, Palmquist DL. Effect of fatty acids or calcium soaps on rumen and total nutrient digestibility of dairy rations. J Dairy Sci 1984;67:978-986.
5. Palmquist DL, Conrad HR. High fat rations for dairy cows: Effects of feed intake, milk and fat production, and plasma metabolite. J Dairy Sci 1978;61:890-901.
6. Palmquist DL, Conrad HR. High fat rations for dairy cows. Tallow and hydrolyzed blended fat at two intakes. J Dairy Sci 1980;63:391-395.
7. Palmquist TC, Jenkins TC. Fat in lactation:Review. J Dairy Sci 1980;63:1-14.
8. Bergen WG, Purcer DB, Cline JH. Effect of ration on the nutritive quality of rumen microbial protein. J Anim Sci 1968;27:1497-1501.
9. AOAC. Official Methods of Analysis.14th ed. Washington (DC): Association of Official Analytical Chemists,1984.
10. Zinn RA, Owens FN. A rapid procedure for purine measurement and its use for estimating net ruminal protein synthesis. Can J Anim Sci 1986;66:157-164.
11. Hill FN, Anderson DL. Comparison of metabolizable energy and productive energy determinations with growing chicks. J Nutr 1958;64:587-594.
12. Zinn RA. Influence of steaming time on site of digestion of flaked corn in steers.J Anim Sci 1990;68:776-781.
13. Goering HK, Van Soest PJ. Forage fiber analyses (Apparatus, reagents, procedures, and some applications) Agric handbook No.379.Washington (DC): ARS-USDA. 1970.
14. Wolin MJ. A theoretical rumen fermentation balance. J Dairy Sci 1960;43:1452-1459.
15. Brouwer E. Report of Subcommitte on Constants and Factors. Procceding of 3rd Symposium on Energy Metabolism;1965 July 14-18; Troon, Scotland. Europe Association of Animal Production Publication. 1965:441-448.
16. National Research Council. Nutrient Requeriments of Beef Cattle. 6th ed. Washington (DC):National Academy Press, 1984.
17. Cochran WG, Cox GM. Experimental design. 2nd ed. New York (NY):Jhon Wiley & Son, 1992.
18. AFOA. Trading and arbitration rules. New York (NY): American Fats and Oil Association, 1988.
19. Grummer RR, Luck ML, Barmore JA. Rumen fermentation and lactation performance of cows fed roasted soybeans and tallow. J Dairy Sci 1993;76:2674-2681.
20. Khorasani GR, de Boer G,Robinson PH, Kennely JJ. Effect of canola fat on ruminal and total tract digestion, plasma hormones, and metabolites in lactating dairy cows. J Dairy Sci 1992;75:492-501.
21. Ohajuruka OA, Wu Z, Palmquist DL.Ruminal metabolism, fiber, and protein digestión by lactating cows fed calcium soap or animal-vegetable fat. J Dairy Sci 1991;74:2601-2609.
22. Elliot JP, Drackley JK,Schauff DJ, Jaster EH.Diets containing high oil corn and tallow for dairy cows during early lactation. J Dairy Sci 1993;76:775-789.
23. Kim YK, Schigoethe DJ, Casper DP, Ludens FC.Supplemental dietary fat from extruded soybeans and calcium soaps of fatty acids for lactating dairy cows. J Dairy Sci 1993;76:197-204.
24. Jenkins TC, Jenny BF. Effect of hydrogenated fat on feed intake, nutrient digestion and lactation performance of dairy cow. J Dairy Sci 1989;72:2316-2324.
25. Drackley JK, LaCount DW, Elliot JP, Klusmeyer TH, Overton TR, Clark JH, et al.Supplemetal fat and nicotinic acid for Holstein cows during an entire lactation. J Dairy Sci 1998;81:201-214.
26. Coppock CE, Wilks DL. Supplemental fat in high-energy rations for lactating cows:Effects on intake, digestion, milk yield, and composition. J Anim Sci 1991; 69:3826-3837.
27. Plascencia A, Zinn RA. Valor nutricional de grasas suplementarias en dietas de crecimiento-finalización para novillos en confinamiento. Vet Méx 1996;27:83-88.
28. Mattos W, Palmquist DL. Increased polynsaturated fatty acids yields in milk of cows fed protected fat. J Dairy Sci 1974;57:1050-1054.
29. Zinn RA, Plascencia A. Comparative digestion of yellow grease and calcium soaps of long-chain fatty acids in cattle. Proceedings of 43th Western Section American Society of Animal Science; 1992; July 8-10; Fort Collins, United States. Fort Collins, United States; American Society of Animal Science,1992:454-457.
30. Zinn RA. Comparative feeding value of supplemental fat in finishing diets for feedlot steers supplemented with and without monensin. J Anim Sci 1988;66;213-227.
31. Zinn RA. Comparative feeding value of supplemental fat in steam-flaked corn and steam-flaked wheat-based finishing diet for feedlot steers. J Anim Sci 1992;70:2959-2969.
32. Doreau M, Legay F, Bauchart D. Effect of source and level of supplemental fat on total and ruminal organic matter and nitrogen digestion in dairy cows. J Dairy Sci 1991;74:2233-2242.
33. Zinn RA. Influence of level and source of dietary fat on its comparative feeding value in finishing diets for feedlot steers:Metabolism. J Anim Sci 1989;67:1038-1049.
34. Zinn RA. Detrimental effect of excesive dietary fat on feedlot cattle growth performance and digestive function. Prof Anim Sci 1994;10:66-72.
35. Elliot JP, Drackley DK, Aldrich CG, Merchen NR. Effect of saturation and sterification of fat sources on site and digestion of organic matter, fiber and nitrogen. J Anim Sci 1997;2803-
36. Zinn RA, Plascencia A. Effects of forage level on the comparative feeding value of supplemental fat in growing-finishing diets for feedlot cattle. J Anim Sci 1996;74:1194-1201.
37. Doreau M, Ferlay A. Effect of dietary lipids on nitrogen metabolism in the rumen:a review. Livestock Prod Sci 1995;43:97-110.
38. Pantoja J, Firkins JL, Eastridge ML.Fatty acids digestibility and lactation performance by dairy cows fed fats varying in degree of saturation. J Dairy Sci 1996;79:429-437.
39. Zinn RA, Alvárez EG, Plascencia A, Shen Y. Influence of method of supplementation on the utilization of supplemental fat by feelot steers. Proceedings of 43th Western Section American Society of Animal Science; 1998; July 27-31; Denver , United States. Denver, United States; American Society of Animal Science,1998:291-296.
40. Palmquist DL, Weisbjerg MR, Hvelplund T. Ruminal, intestinal, and total digestibilities of nutrients in cows fed diets high in fat and undegradable protein. J Dairy Sci 1993;76:1353-1364.
41. NRC. Ruminant Nitrogen Usage. Washington (DC): National Academy Press, 1985.
42. Zinn RA, Bull LS, Hemkin RW. Degradation of supplemental proteins in the rumen. J Anim Sci 1981;52:857-866.
43. Doreau M, Ferlay A, Elmeddah Y. Organic matter and nitrogen digestion by dairy cows fed calcium salts of rapeseed oil fatty acids or rapeseed oil. J Dairy Sci 1993;71:499-504.
44. Christensen RA, Overton TR, Clark JH, Drackley JK, Nelson DR, Blum SA. Effects of dietary fat with or without niconic acid on nutrient flow to the duodenum of dairy cows. J Dairy Sci 1996;79:1410-1424.
45. Klusmeyer TH, Lynchet GL, Clark JH, Nelson DR. Effect of amount of forage and calcium salts of long chain fatty acids(Ca-LCFA) on ruminal fermentation and flow of nutrients to the small intestine of lactating dairy cows. J Dairy Sci 1989;72(Suppl 1):482 (Abstr).
46. Sutton JD, Knight R, McAllan AB, Smith RH. Digestion and synthesis in the rumen of sheep givens diets supplemented with free and protected oils. Br J Nutr 1983;49:419-427.
47. Jenkins TC. Fotouhi N. Effects of lecithin and corn oil on site of digestion, ruminal fermentation and microbial protein synthesis in sheep. J Anim Sci 1990;68:460-466.
48. Plascencia A, Alvárez E, Zinn RA. Efecto de grasa suplementaria y lecitina sobre digestion de nutrientes y fermentación ruminal en dieats para cabras lactantes. Rev Cs Agrop 1991;3:49-58.
49. Czerkawsky J.W. Effect of linseed oil fatty acids and linseed oil on rumen fermentation in sheep. J Agric Sci Camb 1973;81:517-531.
50. Tamminga S, Van Vuuren AM, Van Der Koelen CJ, Khattab HM, Van Gils LGM. Further studies on the effect of fat supplementation of concentrates fed to lactating cows. 3 Effect on rumen fermentation and site of digestion of dietary components, Neth J Agric Sci 1983;31:249-255.
51. Towne G, Nagaraja TG, Brandt RT Jr, Kemp KE. Ruminal ciliated protozoa in cattle fed finishing diets with or without supplemental fat. J Anim Sci 1990.68:2150-2155.
52. NRC, Nutrient Requeriments of dairy Cattle. 6th ed. Washington (DC):National Acedemy Press, 1988.
53. Zinn RA. Influence of level and source of dietary fat on its comparative feeding value in finishing diets for steers: Feedlot growth and performance. J Anim Sci 1989;67:1029-1037.
54. Ferlay A. Doreau M. Influence of method of administration of rapeseed oil in dairy cows. 1. Digestion of nonlipid components. J Dairy Sci 1992;75:3020-3027.
55. Hussein HS, Merchen NR, Fahey CG Jr. Effects of forage level and canola seed supplementation on site and extent of digestion of organic matter, carbohydrates, and energy by steers. J Anim Sci 1995;73:2458-2468.
56. Palmquist DL. Influence of source and amount of dietary fat on digestibility in lactating cows. J Dairy Sci 1991;74:1354-1360.
57. Aldrich G.C., Merchen N.R. Drackley J.K. The effect of roasting temperature applied to whole soybeans on site of digestion by steers:I. Organic matter, energy, fiber, and fatty acids digestion. J Anim Sci 1995;73:2120-2130.
58. Markus SB, Wittenberg KM, Ingalls JR, Undi M. Production response by early lactation cows to whole sunflower seed or tallow supplementation of a diet based on barley. J Dairy Sci 1996;79:1817-1825.
59. Jenkins TC. Butylsoyamide protects soybean oil from ruminal biohydrogenation:Effects of butylsoyamide on plasma fatty acids and nutrient digestion in sheep. J Anim Sci 1995;73:818-823.
60. Tackett VL, Bertrand JA, Jenkins TC, Pardue FE, Grimes LW. Interaction of dietary fat and acid detergent fiber diets of lactating dairy cows.J Dairy Sci 1996;79:270-275.













.jpg&w=3840&q=75)