Nutrición con Aminoácidos en Ganadería de Carne
Publicado: 1 de enero de 2002
Por: Dr. David S. Parker - Novus Europe s.a./n.v.
El propósito de este trabajo es identificar desarrollos en formulación de alimento para rumiantes, los cuales han resultado en un alejamiento de los métodos basados en el cálculo de fibra cruda al sistema, que expresan los requerimientos proteicos sobre la base de proteína absorbida o metabolizable. Este acercamiento ha sido adoptado por un número de planes utilizados en la nutrición de ganado lechero (por ejemplo AFRC en Reino Unido, INRA en Francia y CPM Dairy en Estados Unidos). En estos métodos, los requerimientos de Nitrógeno del rumen para la síntesis de proteína microbiana, son definidos por la energía disponible para la fermentación, y la proteína adicional requerida se logra como fuentes de proteína desviada en el alimento. En novillos de cría, la importancia de lograr los requerimientos del rumen por Nitrógeno ha sido demostrada en estudios donde la suma de proteína degradable en rumen a las dietas de forraje aumentaba el consumo y mejoraba la producción. Este proceso de “sincronizar” el Nitrógeno con la liberación de energía dentro del rumen para mejorar la producción de proteìna microbiana, es estimado como un factor clave para mejorar la eficiencia en la utilización del Nitrógeno por los rumiantes.
En suma a la necesidad de optimizar el funcionamiento del rumen, también se reconoce que el perfil del aminoácido de la proteína que ingresa al duodeno, una mezcla de proteína microbiana y de comida no degradada, puede no ser ideal para una síntesis eficiente de músculo y leche. Experimentos llevados a cabo con animales en crecimiento, han mostrado que, en muchas situaciones, hay respuestas de crecimiento a una provisión aumentada de aminoácidos limitantes –en primer lugar lisina y metionina. La determinación de los “requerimientos” para estos dos aminoácidos provee una base para formular raciones, que optimicen el suministro de aminoácidos y el desarrollo de sistemas de racionado, tales como el NRC (Requerimientos Nutritivos para el Ganado de Carne) de 1996 (Actualizado en el 2000) que toma esto en cuenta. En la mayoría de las situaciones prácticas, este proceso involucra la provisión de materias primas en la ración, el cual balancea el suministro de proteína metabolizable para los aminoácidos limitantes. En la mayor parte de los casos, esto aumentará el suministro completo de proteína por encima de los requerimientos, tal como todas las fuentes de proteína llevarán aminoácidos adicionales al grupo limitante.
Un acercamiento alternativo es el uso de suplementos individuales de aminoácidos “protegidos”, lo que permite al nutricionista llegar a las deficiencias de aminoácidos clave en el suministro de proteína metabolizable, sin aumentar el contenido global de proteína en la dieta. Actualmente, esta opción está restringida al uso de metionina protegida, lo cual ha sido demostrado que mejora el rendimiento del ganado de carne, en un número de pruebas de producción.
1.0 INTRODUCCION
La formulación de raciones para alcanzar los requerimientos de proteínas y aminoácidos del ganado rumiante, ha sido desarrollada de sistemas que estaban basados en proteína cruda digerible (ARC, 1980; NRC, 1984), a aquellos que comenzaron a expresar los requerimientos proteicos en términos de proteína metabolizable o absorbida (MP). Estos métodos tienen en cuenta el impacto de la fermentación en rumen de los componentes proteicos en la dieta, y establecen alcanzar los requerimientos de Nitrógeno para la síntesis de proteína microbiana dentro del rumen. En suma, la necesidad de proteína desviada en rumen en animales con una gran demanda para crecimiento o lactancia, también se ha calculado (AFRC, 1993; NRC, 1996). Los sistemas de racionado de este tipo requieren un conocimiento detallado de las características del alimento, y la aplicación de este acercamiento ha resultado en esfuerzos considerables para correlacionar las características químicas de las materias primas del alimento con fermentabilidad en el rumen, y las predicciones del potencial desviado. Los aspectos clave de la determinación de los valores de MP (proteína metabolizable) para los componentes del alimento, pueden ser definidos como: a) La contribución de la proteína en alimento a la proteína microbiana digerible real. Y b) La contribución de la fuente de proteína al suministro de proteína digerible no degradada. Los elementos esenciales de estos sistemas proteicos están resumidos debajo en la Tabla 1:
Tabla 1: Determinación de valores de Proteína Metabolizable para Alimentos
Componente | Factores que afectan el Valor del Alimento |
| Proteína Degradable en Rumen | Solubilidad – Fracción Rápidamente Degradada/Lentamente Degradada Índice de Secreción del Rumen Nivel de Alimentación |
| Síntesis de Proteína Microbiana | Suministro de Energía Dentro del Rumen Suministro de Nitrógeno Dentro del Rumen |
| Proteína Digerible No Degradada | Alcance de la Degradabilidad del Rumen Proporción de Nitrógeno Insoluble (por ej.: Lignina Ligada) Composición de Aminoácido del Material desviado |
| Uso de Aminoácidos | Eficiencia de Absorción Eficiencia de Utilización para Mantenimiento, Crecimiento, Lactancia, Preñez |
2.0 TRANSACCION DE NITROGENO EN RUMEN
2.1 Requerimientos para la síntesis de proteína microbiana:
El suministro de proteína microbiana al duodeno puede alcanzar entre el 50 y el 90% de los requerimientos de MP (Proteína Metabolizable) del ganado de carne, y un elemento clave de los sistemas modernos de formulación de raciones es la optimización del suministro de nutrientes en el rumen para maximizar este proceso. El crecimiento de proteína microbiana está enlazado con la energía disponible dentro del rumen de la fermentación. Las asignaciones están hechas para el contenido en grasas y los productos finales fermentables presentes en forrajes ensilados, en ambos sistemas (ARC e INRA). El sistema NRC utiliza la digestión de carbohidratos en el rumen como pronosticador de la síntesis de proteína microbiana, a pesar de que hay pocos datos disponibles para la digestión e índices de pasaje en el ganado. Un resumen de valores de la literatura muestra que la síntesis de proteína microbiana está entre 12,6 y 17 g/100 g TDN en la ración.
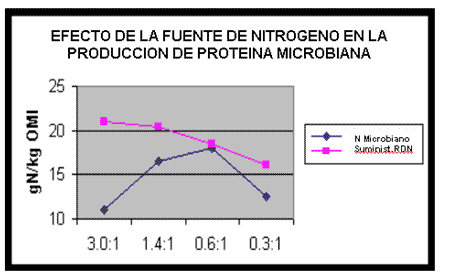
El impacto de la fuente de Nitrógeno disponible a la población microbiana en la eficiencia global de la producción proteica ha sido también investigado. El efecto de cambiar el radio de urea a harina de pescado en un suplemento isonitrogenado para alimentar al ganado de carne, con una dieta basada en silaje de maíz sobre la síntesis de proteína microbiana, se muestra en la Figura 1 (Cotterill et al, 1982).
Puede verse que en los índices más altos y más bajos de inclusión de urea, la producción de proteína microbiana fue limitada. La síntesis microbiana aumentó con la proporción incrementada de harina de pescado, aunque el suministro global de Nitrógeno degradable en rumen (RDN) fue reducido. Estudios de este tipo demuestran la importancia de una provisión de proteína de consumo degradable (DIP) para estimular la síntesis de proteína microbiana.
2.2 Efecto sobre el consumo y la digestibilidad:
El impacto de la fuente de DIP (proteína de consumo degradable) en la estimulación de la fermentación microbiana y en el aumento del consumo de forrajes de calidad pobre, está bien establecido. Datos de Köster et al (1996) mostraron que incrementar los niveles de DIP en las dietas basadas en forraje de pastos altos de pradera, de 0 a 720 g/d, mejoró la eficiencia de producción de proteína microbiana de 12,2 a 20,0 gN/kg OM verdaderamente digerida en el rumen. En un estudio similar reportado por Mathis et al (1999), la adición de suplementos de poroto de soya para forrajes de baja calidad, aumentó el consumo de forraje y el rendimiento global del ganado de carne. Estas mejoras fueron asociadas con la fermentación aumentada en el rumen y un incremento en los niveles de Amoníaco en el rumen, indicando que en dietas con forrajes de baja calidad, el Nitrógeno soluble estuvo limitando la producción microbiana. En esta situación, la suplementación con proteína de consumo no degradada (UIP), mientras se incrementa el suministro duodenal de proteína, no mejoraría el consumo y la digestibilidad del material de forraje debido a limitaciones impuestas sobre la actividad microbiana por una baja disponibilidad de Nitrógeno en el rumen. En un experimento reciente de Bodine et al (2000), la relación entre el suministro de DIP (proteína de consumo degradable), y el consumo y la digestibilidad de heno de pradera de baja calidad, fue examinada. La escasa digestibilidad de materia orgánica de heno de pradera (0,3) en raciones conteniendo maíz enrollado, fue significativamente aumentada a 0,5 cuando el requerimiento de DIP del rumen fue alcanzado utilizando harina de soya. Otro trabajo de Bandyk et al (2001), investigó el impacto del suministro aumentado de proteína al rumen y al duodeno, en el consumo y digestibilidad de forraje de baja calidad. Los resultados mostraron que, mientras ambas fuentes mejoraban el consumo, el suministro de DIP al rumen fue el más efectivo, estimulando el consumo de materia orgánica en un 65%.
2.3 Mejorando la captura de Nitrógeno:
La importancia de balancear la disponibilidad de energía y nitrógeno dentro del rumen, y su efecto en la eficiencia de la síntesis de proteína microbiana ha sido investigada en cierto número de estudios. La “eficiencia” de captura del Nitrógeno soluble presente en forrajes frescos ensilados es baja, y puede ser mejorada por el uso de suplementación apropiada. En los estudios de Rooke et al (1987), las infusiones intrarruminales de Nitrógeno soluble adicional en animales alimentados con silaje de pasturas, no tuvieron efecto sobre la eficiencia de crecimiento de proteína microbiana. La provisión de una fuente de energía fácilmente fermentable (jarabe de glucosa), sin embargo, incrementó el flujo de Nitrógeno no Amoníaco (NAN) al duodeno, y redujo significativamente la concentración de Amoníaco en el rumen. En estudios más recientes, llevados a cabo en el Instituto para la Investigación Ambiental y de Pastizales en el Reino Unido, el impacto del uso de forrajes seleccionados por su alto contenido de carbohidratos solubles en agua (WSC) para la captura de Nitrógeno, ha sido investigado. En un experimento con corderos en cría, Lee et al (1999) reportó que las pasturas para pastoreo conteniendo hierbas con alto nivel de WSC (carbohidratos solubles en agua), resultaron en una mejora significativa en la ganancia de peso, cuando se las comparaba con una variedad baja en WSC. El contenido de Nitrógeno de ambas variedades era similar. Otro trabajo del Grupo IGER (Miller et al, 1999) con vacas lecheras pastando ambas variedades, mostró que los animales alimentados con la variedad con WSC alto, dividió una proporción más alta de consumo de Nitrógeno en leche. Este estudio subraya el potencial para incrementar la eficiencia en la captura de Nitrógeno soluble de los forrajes, igualando la liberación de energía y la disponibilidad de Nitrógeno. A pesar del interés en este tema, no hay una clara indicación para precisar la relación entre los índices de liberación de energía y de Nitrógeno, y la eficiencia del rumen en conjunto (para repasar, ver Sauvant y van Milgen, 1995), lo que puede ser llevado a la práctica fácilmente.
3.0 ALCANZANDO LOS REQUERIMIENTOS DE AMINOACIDOS
3.1 Contribución de proteína de consumo no degradada:
A pesar de balancear la dieta para mejorar la eficiencia en la captura de Nitrógeno en el rumen, y por lo tanto, el flujo de proteína microbiana al duodeno mejoraría el rendimiento del animal, se necesita UIP (proteína de consumo no degradada) adicional para alcanzar los requerimientos de proteína metabolizable en animal de gran rinde. Los novillos en crecimiento alimentados con una ración de forraje/grano, formulada para llegar a los requerimientos de Nitrógeno del rumen y el crecimiento de 1 kg/d, mostraron índices mejorados de ganancia seguidos de la infusión de cantidades clasificadas de caseína en el duodeno (Beerman et al, 1997). Otras pruebas, reportadas por el mismo Grupo, en las cuales la infusión de caseína fue reemplazada por niveles en aumento de UIP (proteína de consumo no degradada) de una mezcla de proteínas animales, confirmaron que el suministro suplementario de proteína al duodeno fue necesario para optimizar el rinde de crecimiento. Similarmente, una serie de pruebas de alimento reportadas por Kunkle y Hopkins (1999a) mostraron la relación entre el consumo suplementario de UIP y la ganancia adicional en ocho estudios llevados a cabo en Florida. La mejora global fue calculada en 140 g/d comparada con los grupos de control donde los requerimientos de proteína degradable en rumen habían sido alcanzados. Está claro por esos estudios que la respuesta a las fuentes de UIP es variable, y depende de la habilidad del balance de aminoácidos en el flujo proteico al duodeno, para llegar a los requerimientos para el crecimiento. Una evaluación de la relación entre la composición de aminoácidos de la bacteria del rumen, la proteína duodenal y los tejidos de la carcaza, se muestran en la Tabla 2.
Tabla 2: Composición de Aminoácidos de Diferentes Fuentes de Proteínas (%)
Metionina Lisina Histidina Fenilalanina Treonina Leucina Isoleucina Valina Arginina Cisteína
a. Wilkerson et al (1993); (b) Klemesrud et al (2000c); (c)Rohr & Lebzien (1991); (d) Ainslie et al (1993)
Es aparente de estas figuras que, aunque la proteína microbiana es el contribuyente principal en el flujo de proteínas en el duodeno, la contribución de UIP (proteína de consumo no degradada) dietaria influencia el patrón global de los aminoácidos presentes. Es también importante reconocer que estas figuras representan valores medios para situaciones específicas experimentales, y habrá una variación considerable, aún en la composición de proteína microbiana, la cual influenciará la habilidad del suministro de proteína metabolizable para alcanzar los requerimientos del animal. (Clark et al, 1992). En NRC 1996, por ejemplo, la contribución de las paredes celulares y no celulares de proteína bacteriana es calculada, cada fracción utilizando una composición de aminoácido diferente. El impacto de niveles aumentados de aminoácidos desviados de diferentes proteínas de alimento sobre la ganancia y los niveles de aminoácidos en sangre (Gibb et al, 1992), demuestra la sensibilidad del sistema. En estas experiencias, la adición de grano seco de destilería a un suplemento de urea, resultó en un aumento de en la ganancia de peso y de metionina en plasma, con el consumo de granos de destilería. Las respuestas sugirieron una deficiencia marginal en el suministro de metionina a bajos niveles en la adición de granos a la dieta seguida por la lisina, que se volvió el aminoácido limitante en altos niveles de inclusión.
En dietas en las que la contribución de UIP (proteína de consumo no degradada), proviene primitivamente de una sola fuente, las deficiencias en aminoácidos específicos y el impacto en el rinde del animal pueden ser identificados. Experimentos reportados por Klopfenstein et al (1995) muestran que, cuando el maíz es la fuente principal de MP (proteína metabolizable) para el ganado en crecimiento, el uso de granos dañados por el calor, en los que la disponibilidad de lisina es reducida, resulta en un índice bajo de ganancia sobre los controles de alimentación con urea, cuando se lo compara con la alimentación con granos no dañados. Este y otros datos de animales alimentados con forraje suplementado con maíz, indican que la lisina es el primer aminoácido limitante. Similarmente, el rendimiento fue realzado en dietas conteniendo harina de carne y hueso, los que son conocidos por su deficiencia en metionina, por la adición de aminoácido protegido.
3.2 Definiendo los requerimientos de Aminoácidos:
Los requerimientos de aminoácidos de los animales en crecimiento han sido estimados para el mantenimiento y la producción, utilizando ambos un acercamiento factorial, y también por medición directa. Un resumen de los datos de los experimentos usando el acercamiento factorial (Williams, 1994), subraya el problema de este método, particularmente la falta de estimados de los requerimientos de mantenimiento de los animales en crecimiento, y los valores asignados a la eficiencia en la utilización de aminoácidos absorbidos por distintas funciones. Históricamente, la aproximación de medición directa ha estado basada en las respuestas del animal a niveles evaluados de suplementación de los aminoácidos apuntados, a menudo implantados directamente en el abomaso de los animales alimentados con una dieta baja en proteínas o semi-purificada. En estudios de este tipo, los cambios en la concentración en sangre de aminoácidos son utilizados para identificar el punto al cual el requerimiento es alcanzado, y entonces excedido. Este “punto de quiebre” en la concentración en sangre es interpretado como el punto al cual el requerimiento es excedido, y los aminoácidos apuntados se acumulan. Usando esta aproximación, Fenderson y Bergen (1975) identificaron metionina como el primer aminoácido limitante, sin indicación clara del segundo. Otro trabajo de Richardson y Hatfield (1978) confirmó la metionina como al aminoácido limitante para la ganancia en animales alimentados con dietas semi-purificadas, con una dieta de 9,3% de proteína cruda. Las infusiones de lisina, treonina y triptofano no mostraron un “punto de quiebre” claro, sugiriendo lo que debe actuar como segundo aminoácido limitante, una vez que el requerimiento de metionina ha sido alcanzado. En una extensión de este “punto de quiebre”, las mediciones aproximadas de retención de Nitrógeno han sido utilizadas para cotejar cambios en la disponibilidad de aminoácido, con aumentos en la acumulación de proteína. Usando esta metodología, Titgemeyer y Merchen (1990a y 1990b) mostraron que, para una dieta base suplementada con una infusión de aminoácidos no conteniendo ni metionina ni cisteína, los requerimientos de aminoácido sulfurosos para novillos de 325 kg de peso, ganando 1,3 kg/día, fue de 14,4 g/d. En este caso, el requerimiento pudo sustancialmente ser alcanzado de la producción de proteína microbiana, y tanto los niveles en sangre de metionina como los datos de retención de Nitrógeno, indicaron una máxima eficiencia en la deposición de proteína cuando un adicional de 3 g/d de metionina fue suministrado a los animales. En otro estudio de Campbell et al (1997), una aproximación similar fue adoptada para evaluar los requerimientos de metionina para novillos de 160 kg, bajo condiciones en las cuales los requerimientos de cisteína fueron excedidos, o la cisteína fue limitada. En este caso, el análisis del “punto de quiebre” de la respuesta a la retención de Nitrógeno indicó un nivel más alto de requerimiento (7.9g/d) que aquel determinado por cambios en los valores de metionina en plasma. Los autores sugieren que las mediciones simples de los niveles de aminoácido en sangre puede ser un pobre reflejo de los cambios en el flujo entre la baja concentración de aminoácidos individuales en sangre y los “charcos” más extensos dentro de los tejidos. La respuesta cuadrática para los aminoácidos totales en sangre en este estudio indica que hubo una respuesta en dos fases al suministro adicional de metionina, el segundo “punto de quiebre” indica alcanzar los requerimientos actuales. Cálculos similares de Froidmont et al (2000) para toros Belgas Azules de musculatura doble en crecimiento, resultaron en un requerimiento de metionina de 26 g/d basado en las mediciones de retención de Nitrógeno. Esto representa 330 mg de metionina/gNitrógeno retenido, lo cual es más alto que los cálculos de otros experimentos (226mg/gN retenido Campbell et al (1997); 279mg/gN retenido Titgemeyer y Merchen (1990b)). Los autores atribuyen esto a la alta retención de Nitrógeno y el potencial de crecimiento de esta raza en particular.
3.3 El uso de Aminoácidos protegidos:
La disponibilidad de métodos de protección en rumen para nutrientes específicos como los aminoácidos, ha suministrado una oportunidad para el balance de alimentos para rumiantes, para alcanzar los requerimientos para los primeros aminoácidos limitantes. El impacto del uso de estos productos en el rendimiento, ha sido variable, reflejando nuestra falta de conocimiento de los requerimientos precisos de aminoácidos específicos, y la dificultad para predecir el flujo de nutrientes individuales al duodeno. Trabajos tempranos con suplementación de raciones basadas en silajes de maíz/maíz/soya, con metionina/lisina protegidas en rumen, resultaron en mejoras significativas de ganancia en animales jóvenes, aunque esto no fue aparente en animales mayores, donde la soya fue reemplazada por urea (Oke et al, 1986). En éste y otro estudio similar por Wright y Loerch (1988), la suma de material protegido resultó en un aumento en los niveles de metionina en sangre, y los tratamientos donde hubo respuesta sin aumento en los niveles de lisina en sangre, sugieren que la lisina fue el aminoácido limitante para el crecimiento. En otro estudio de Hussein y Berger (1995), utilizando una dieta con un 71 % de maíz descascarado/silaje de maíz, suplementado con soya o con soya/urea, no hubo un beneficio claro por la adición de metionina/lisina protegida en rumen. Estos autores especularon que el bajo pH del rumen asociado con una degradabilidad más baja de las fuentes de proteína dietaria, reduciendo la necesidad de aminoácidos suplementarios.
En una serie de experiencias investigando el impacto del balance del suministro global de aminoácidos, el Grupo de Nebraska ha desarrollado un acercamiento interesante utilizando fuentes simples de suplementos proteicos. En un estudio con harinas de carne y hueso agregada a la ración base de silaje de sorgo y mazorcas de maíz, Klemesrud et al (1997) mostró que la adición de metionina protegida resultó en una mejora significativa en el rendimiento. La suma de una fuente de lisina protegida no tuvo efecto, confirmando que la metionina fue el primer aminoácido limitante en esas raciones. Otra experiencia con harina de gluten de maíz (Klemesrud et al, 2000a) demostró una respuesta con lisina protegida, y este efecto fue confirmado con una dieta basada en silaje de maíz suplementado con gluten de maíz (Klemesrud et al, 2000b). En ambos experimentos, las concentraciones en sangre del aminoácido limitante quedaron constantes, reflejando el uso de aminoácidos adicionales de desvío para la ganancia. La importancia de balancear las fuentes de proteína, y la relación entre el suministro total de aminoácidos sulfurosos y la proporción de metionina/cisteína, fue investigada (Klemesrud et al, 2000c) usando suplementos de harinas de carne y hueso (deficientes en metionina) y harina de plumas (alto contenido de aminoácidos sulfurosos, pero mayormente como cisteína). Aunque la suma de harina de plumas mejoró el rendimiento de los animales alimentados con harinas de carne y hueso, sólo la adición de metionina protegida en rumen probó ser el mejor tratamiento, indicando que el balance de metionina a cisteína había sido rectificado. Los autores reportan un efecto similar cuando fue agregada metionina protegida a la ración que contenía harinas de sangre y de plumas. Una revisión del impacto de suplementación del forraje en animales alimentados con distintas fuentes de metionina, como el consumo de proteína no degradada o aminoácidos protegidos en rumen, por Kunkle y Hopkins (1999b), mostró un incremento lineal en ganancia de peso al agregar a la dieta hasta unos 5g/d de aminoácidos de desvío sulfurosos. La respuesta fue similar para las fuentes de proteína no degradada anexadas como para la metionina protegida, indicando que, en esta situación, la metionina fue el primer aminoácido limitante. En una experiencia separada en la cual se utilizaron diferentes fuentes de metionina protegida, (Kunkle, comunicación personal; Davis et al, 2000), fue notado que no todos los productos daban el mismo nivel de rendimiento, reflejando posiblemente distintos grados de protección dentro del rumen.
4.0 CONCLUSION
El desarrollo de nuestras ideas concernientes a la formulación de raciones para especies rumiantes, es proveer oportunidades para optimizar el funcionamiento del rumen, y alcanzar los requerimientos del animal para proteína metabolizable. En las etapas iniciales, este proceso apunta a ligar la energía disponible de la fermentación a la provisión de nitrógeno, para maximizar la producción de proteína microbiana. También es claro que la sincronización mejorada de este proceso beneficiará la economía de nitrógeno en conjunto del animal, y que las fuentes de proteína degradable tiene un rol importante que jugar en estimular la digestión y la ingesta de forraje. Aunque la proteína microbiana tiene un alto valor biológico, se considera generalmente que la metionina es el primer aminoácido limitante en la proteína, y se confirma por medio de estudios, donde la metionina adicional de desvío ha sido provista. En situaciones de alimentación, donde un monto significativo de proteínas de desvío del rumen ingresan al duodeno, esto impactará sobre la composición de aminoácidos de la provisión de proteína metabolizable en el animal, y podrá resultar en la transformación de un aminoácido particular limitante para una eficiente deposición de tejidos. Experimentalmente, esto ha sido demostrado para alimento con gluten de maíz (la lisina se transforma en aminoácido limitante), y para alimento de carne y hueso (la metionina se transforma en aminoácido limitante). También ha habido especulación en que el triptofano y la histidina pueden tener un rendimiento limitante en ciertas circunstancias. En términos prácticos, la única fuente comercialmente disponible de aminoácidos simples es la metionina, y es posible demostrar un beneficio financiero para su uso en situaciones particulares. En ausencia de algún desarrollo técnico, el cual proveerá suplementos de aminoácidos protegidos, el uso de programas tales como CNCPS proveerá un mecanismo para alcanzar la proteína metabolizable y los requerimientos de aminoácidos del animal.
Referencias:
AFRC (1993) Energy and protein requirements of ruminants. CAB, UK
Ainslie S.J., Fox D.G., Perry T.C., Ketchen D.D. and Barry M.C. (1993) Journal of Animal Science 71, 1312-1319
ARC (1980) The nutrient requirements of ruminant livestock. CAB, UK
Bandyk C.A., Cochran R.C., Wickersham T.A., Titgemeyer E.C., Farmer C.G. and Higgins J.J. (2001) Journal of Animal Science 79, 225-231
Beermann D.H., Robinson T.F., Knaus W.F. and Fox D.G. (1997) Proceedings of the Cornell Nutrition Conference, October 1997
Bodine T.N., Purvis H.T., Ackerman C.J. and Goad C.L. (2000) Journal of Animal Science 78, 3144-3154
Campbell C.G., Titgemeyer E.C. and St-Jean G. (1997) Journal of Animal Science 75, 230-238
Clark J.H., Klusmeyer T.H. and Cameron M.R. (1992) Journal of Dairy Science 75, 2304
Cotterill B.R., Beever D.E., Austin A.R. and Osbourn D.F. (1982) British Journal of Nutrition 48, 527-541
Davis L.B., Kunkle W.E., Bates D.B. and Reiling B.A. (2000) Journal of Animal Science 78 Suppl. 1, 116-117
Fenderson C.L. and Bergen W.G. (1975) Journal of Animal Science 41, 1759-1766
Froidmont E., Beckers Y and Thewis A. (2000) Journal of Animal Science 78, 233-241
Gibb D.J., Klopfenstein T.J., Britton R.A. and Lewis A.J. (1992) Journal of Animal Science 70, 2885-2892
Hussein H.S. and Berger L.L. (1995) Journal of Animal Science 73, 3503-3509
Klemesrud M.J., Klopfenstein T.J. and Lewis A.J. (1997) Journal of Animal Science 75, 3301-3306
Klemesrud M.J., Klopfenstein T.J. and Lewis A.J. (2000a) Journal of Animal Science 78, 199-206
Klemesrud M.J., Klopfenstein T.J. and Lewis A.J. (2000b) Journal of Animal Science 78, 207-215
Klemesrud M.J. , Klopfenstein T.J., Stock R.A., Lewis A.J. and Herold D.W. (2000c) Journal of
Animal Science 78, 1060-1066
Klopfenstein T.J., Klemesrud M.J., Herold D.W., Stock R.A. and Lewis A.J. (1995) Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, 67-75
Köster H.H., Cochran R.C., Titgemeyer E.C., Vanzant E.S., Abdelgadir I. And St-Jean G. (1996) Journal of Animal Science 74, 2473-2481
Kunkle W.E. and Hopkins D.I. (1999a) Feedstuffs Feb. 15th, 11
Kunkle W.E. and Hopkins D.I. (1999b) Proceedings of the 10th Florida Ruminant Nutrition Conference, 28-38
Lee M.R.F., Jones E.L., Humphreys M.O., Moorby J.M., Dhanoa M.S., Theodorou M.K., MacRae J.C. and Scollan N.D. (1999) South African Journal of Animal Science 29, 281
Mathis C.P., Cochran R.C., Stokka G.L., Heldt J.S., Woods B.C. and Olson K.C. (1999) Journal of
Animal Science 77, 3156-3162
Miller L.A., Theodorou M.K., MacRae J.C., Evans R.T., Andesogan A.T., Humphreys M.O., Scollan
N.D. and Moorby J.M. (1999) South African Journal of Animal Science 29, 281
NRC, Nutrient Requirements of Beef Cattle (Update 2000) (1996) National Academic Press, Washington, USA
Oke B.O., Loerch S.C. and Deetz L.E. (1986) Journal of Animal Science 62, 1101-1112
Richardson C.R. and Hatfield E.E. (1978) Journal of Animal Science 46, 740-745
Rohr K. and Lebzein P. (1991) Proceedings of the 6th International Symposium on Protein
Metabolism and Nutrition. EAAP Publication No. 59, 127-134
Rooke J.A., Lee N.H. and Armstrong D.G. (1987) British Journal of Nutrition 57, 89-98
Sauvant D. and van Milgen J. (1995) Proceedings of the 8th International Symposium on Ruminant
Physiology. F.E. Verlag, Stuttgart, Germany, 71-87
Titgemeyer E.C. and Merchen N.R. (1990a) Journal of Animal Science 68, 750-757
Titgemeyer E.C. and Merchen N.R. (1990b) Journal of Animal Science 68, 2075-2083
Wilkerson V.A., Klopfenstein T.J., Britton R.A., Stock R.A. and Miller P.S. (1993) Journal of Animal
Science 71, 2777 – 2784
Williams A.P. (1994) In: Amino Acids In Farm Animal Nutrition. Ed. J.P.F. D’Mello, CABI, UK
Wright M.D. and Loerch S.C. (1988) Journal of Animal Science 66, 2014-2027
Temas relacionados:
Recomendar
Comentar
Compartir

22 de febrero de 2011
Buen dia: Mi nombre es alejandro contreras soy químico farmaceutico , estoy realizando un estudio donde estoy evaluando productos veterinarios conocidos como modificadores orgánicos, por el presente medio quisiera que me colaboraran con alguna información acerca de estos productos envió un poco de la descripción de mi inquietud. Comercialmente se encuentra en el mercado veterinario productos conocidos como "modificadores orgánicos" para bovinos que es utilizado : -Al momento de destete para controlar el estres en los animales -En los meses finales de la ceba para mejorar este proceso. -En hembras para mejorar condición y aumento de defensas En hembras cuando terminan lactancia -Coadyudante en múltiples patologías : Coadyudante en la mejora de problemas reproductivos -coadyudante en el tratamiento de deficiencias de aminoacidos o vitaminas Los modificadores orgánicos están compuestos por: Aminoacidos, vitaminas y oligoelementos y se presentan como soluciones inyectables, de administración subcutánea. Es importante aclarar la composición de estos productos, por esta razón me comunico con ustedes ya que existe la duda de cuales son las concentraciones EFECTIVAS y PERMITIDAS de aminoacidos en este tipo de producto.? teniendo encuenta que la administración es dosis única y es utilizada en casos de estres en animales adultos (400-600 Kg) y terneros (50-150 Kg), además la mayoría de ganado vacuno es alimentado con forraje el cual es un alimento completo. Esta es la lista con los aminoacidos que contienen estas formulaciones: L-Glu, L-Asp,L-His,L-Ser,L-Gly,L-Thr,L-Ala,L-Arg,L-Tyr,L-Cys,L-Val,L-Met,L-Phe,L-Ile,L-leu,L-Lys,L-Pro. quisiera se me aclara los limites de concentración de estos amino acidos. Cordialmente Alejandro Contreras
Recomendar
Responder


8 de octubre de 2010
quisiera saber los requerimientos nutricionales de los bovinos de carne en sus distintas edades porque se que si el animal es de menor edad necesitaria mas proteina que un animal adulto:
quisiera sabe para poder elaborar una racion alimenticia como sostenimiento de los animales o como para engorde:
que porcentaje de proteina necesitan?
aminoacidos [percent]?
[percent] de grasa?
[percent] de energia?y minerales como calcio, fosforo, potasio u otros escenciales para su engorde?
saludos....
gracias...
Recomendar
Responder
6 de septiembre de 2005
Los rumiantes no necesitan aminoácidos esenciales, debido a que ellos con la flora ruminal obtienen todos los aminoácidos que necesitan, entendiéndose por aminoácido esencial aquel que el animal no puede sintetizar y lo tiene que conseguir en la dieta, por decirlo así.
Recomendar
Responder
18 de agosto de 2005
Existe algun trabajo donde se especifique el gusto que toma la carne luego de producida con distintos alimentos. Estoy para todo, creo que nadie se interesa en lo mÁs importante, es decir el gusto que toma la carne.
Recomendar
Responder
16 de agosto de 2005
Hola, los rumiantes qué tipo de aminoácidos pueden conusmir: esenciales o no esenciales.
Recomendar
Responder
14 de agosto de 2005
Con qué puedo aumentar la proteina de consumo degradada para aumentar 140'g por dia de engorde del animal, y otra pregutnta es cómo le darian la metionina, como lo podria hacer y como. Bueno tal vez a ustedes no les interesa pero a mi me interesa y más que mucho como influye en el gusto de los animales los distintos tipos de alimentación.
Recomendar
Responder
13 de agosto de 2005
En que proporción le darias aminoacidos para aumentrar la producción de carne, cuanto aumentaria, cuánto salen si sabes en canrgfentina, pero bueno, es decir enbsintesis cuanto influenciaria en mi engorde a corral darles esos aminoacidos, cual es su nombre especifico.
Recomendar
Responder


6 de agosto de 2005
Sobre la influencia de los suelos en la producción forrajera es muy importante, es por ello
que se recomienda realizar un análisis de suelos para ver sus componentes mas significativos como el N-P-K, las proporciones de arena, limo y arcilla, su pH y carga eléctrica.
De acuerdo a ello se puede realizar su clasificación y sus requerimientos en la fertilización.
En la nutrición es vital tener presente los niveles de proteína y de NDT, complementados
con las sales minerales, dependiendo sobre el tipo de explotacion que se lleve, ya sea
extensiva, semi-extensiva y estabulada, para una mejor eficiencia nutricional.
Igualmenete la raza de ganado que se esta trabajando, complementada con los rubros
de sanidad, medio ambiente y manejo.
Sobre los niveles de metionina, lisina, triptofano y otros animoácidos están en proporción de los insumos que se utilicen en la alimentación y como referencia hay tablas como la NRC que se actualizan, segun las necesidades nutricionales.
Recomendar
Responder
13 de junio de 2005
Luiz Zerpa Vilela. ing. zootecnista Peru
Cuál es la influencia de los suelos en la produccion forrajera y en
el contenidos de aminoacidos en el ganado vacuno.
Recomendar
Responder
18 de octubre de 2004
en que productos puedo conseguir metionina suplemantaria y lisina.
Recomendar
Responder
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar

Norel Animal Nutrition