Introduccion
La caída sostenida de producción de grasa en leche se traduce en pérdidas económicas significantes en un establo porque el precio de la leche se basa en los componentes en Estados Unidos. Afortunadamente, muchos productores experimentan pocos problemas con la DGL debido a que sus nutricionistas han desarrollado y mantenido un programa de alimentación bien balanceado y consistente. Aún el mejor nutricionista, sin embargo, puede ser víctima de la DGL después responder a cambios en precios de los ingredientes, disponibilidad limitada de algún ingrediente o cambios no esperados en la composición de nutrientes de los ingredientes. Aparentemente los cambios lógicos en el programa de alimentación pueden disminuir la grasa de la leche varias fracciones de un punto de porcentaje a más de un punto porcentual completo en un período de tiempo corto. Esto puede tomar varias semanas o meses para identificar la causa nutricional y regresar a una producción de grasa normal.
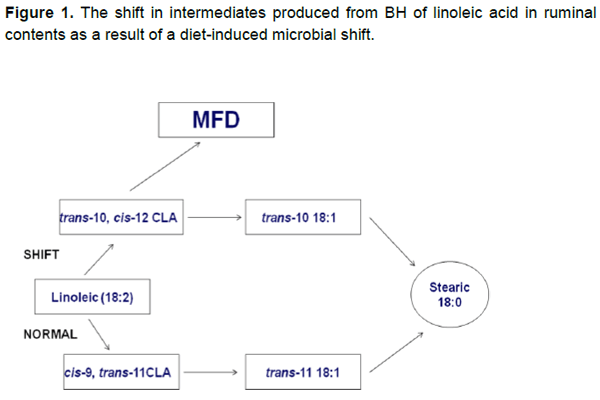
La DGL es causada por cambios conducidos por nutrición en el rumen. Los lípidos en el alimento son metabolizados por la población microbiana del rumen en una vía llamada BH, la cual conduce a la formación de lípidos bioactivos (figura 1). “Bioactivo” significa que los lípidos afectan tejido y células vivas. Estos lípidos bioactivos son referidos como ALC, siendo el más común el ALC cis-9, trans-11 (Figura 1). Los microorganismos en el rumen producen más de veinte tipos de ALC conocidos, pero se ha demostrado que tres de ellos causan DGL. Esta discusión se referirá a estos tres como ALCIGL debido a que estos ALC actúan como inhibidores de la grasa de la leche. El más estudiado de los tres ALCIGL es el ALC trans-10, cis-12. Los ALCIGL producidos en el rumen viajan vía sanguínea a la glándula mamaria, donde inhiben la síntesis de grasa de la leche mediante el daño de la producción de varias enzimas esenciales para la síntesis de grasa en la glándula mamaria (Bauman y Griinari, 2003). Los ALCIGL están también presentes en vacas que producen niveles aceptables de grasa en leche, pero en concentraciones muy pequeñas para causar DGL.
Finalmente, el tipo de alimento que la vaca consume afecta las condiciones del rumen, el cual a su vez afecta la cantidad y tipo de ALC producido. Considerando que la sobreproducción de ALCIGL en el rumen conduce a DGL, el exceso de ALCIGL y consecuentemente la DGL pueden ser controladas mediante la observación de cerca de varios riesgos nutricionales. Este artículo engloba estos riesgos y así otorga control al nutricionista para la síntesis de grasa en la leche.
CAMBIO EN EL PERFIL DE LOS ÁCIDOS GRASOS EN LECHE DURANTE LA DGL
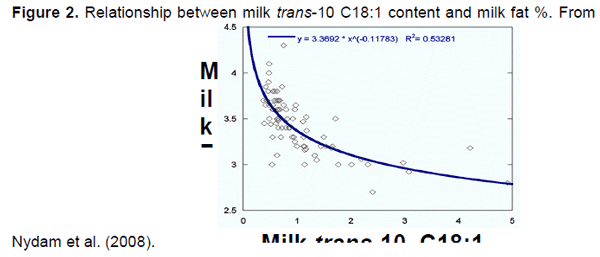
Los cambios en el perfil de ácidos grasos pueden ser observados en la leche conforme el porcentaje de grasa varía. Un ácido graso en leche que se correlaciona bien con la grasa de la leche es el intermediario BH trans-10 C18:1 (Figura 2). Conforme la concentración de trans-10 C18:1 en leche incrementa, se presenta una disminución curvilínea en el porcentaje de grasa en la leche. El análisis de leche para trans-10 C18:1 predice bien el porcentaje de grasa en leche, aunque el porcentaje de grasa real es bien conocido con antelación para revisar los reportes de laboratorio anteriores de ácidos grasos en leche. Una elevación de trans-10 C18:1 en leche confirma que la vía ruminal de BH ha sido alterada, apuntando a una causa nutricional de la baja de grasa en leche.
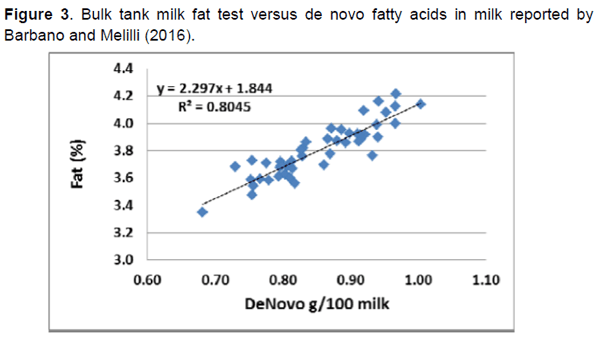
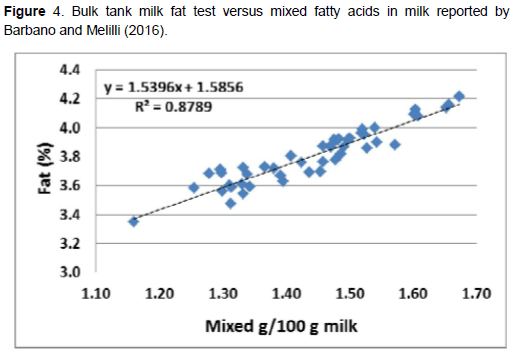
Más recientemente, las técnicas de espectroscopia media-infrarroja han sido utilizada para un análisis rápido de ácidos grasos, incluyendo ácidos grasos de novo (C4 a C14), origen mixto (C16:0, c16:1 y C17:0, y preformados (C18 y de cadena más larga). Los ácidos grasos de novo incrementaron linealmente (Figura 3) conforme incrementó el porcentaje de grasa en la leche en una serie de datos de análisis media-infrarroja de muestras de leche de 39 establos. Los ácidos grasos en leche fueron clasificados en 19 establos como altos en ácidos grasos de novo (promedio 23.10 g/100 de ácidos grasos). También, los ácidos grasos mixtos en leche incrementaron con la grasa en la leche (Figura 4) pero no hubo relación de ácidos grasos preformados con la grasa en leche. Barbano y Melilli (2016) concluyeron que los establos necesitaban tener una concentración de ácidos grasos de novo en al menos 0.85 g/100 g de leche para alcanzar una prueba de grasa en tanque mayor a 3.75%. Un estudio realizado por Woolpert et al. (2016) indicó que la sobrepoblación, redujo la frecuencia de alimentación y una mayor grasa dietética estuvo asociada con una menor producción de ácidos grasos de novo en leche. En un estudio posterior, Woolpert et al. (2017) report que el manejo de la alimentación, la densidad animal, la grasa dietética y las características físicas de la dieta, todos son relacionados con los ácidos grasos de novo y pueden ser factores de riesgo para la depresión de grasa en leche.
El uso de ácidos grasos en leche continúa creciendo como un medio para monitorear un número de parámetros de producción y salud de las vacas. Dorea et al. (2017) mostró que los ácidos grasos en la leche están relacionados a las concentraciones de AGNE en el plasma y podrían ser utilizados como un indicador del balance de energía del animal. Las mejores correlaciones de AGNE en plasma fueron observadas para la tasa de ácidos grasos en leche de C18:0 a C15:0, y C17:0 a C15:0. En otra aplicación, Coppa et al. (2017) utilizaron el análisis espectroscopia medio-infrarroja en leche para desarrollar ecuaciones para predecir la composición de ácidos grasos en leche y su valor potencial para el beneficio de la salud humana.
FACTORES NUTRICIONALES QUE INFLUENCÍAN LA GRASA EN LA LECHE
Los factores nutricionales que tienen influencia sobre el contenido de grasa en leche pueden ser divididos en principales conductores y sintonizadores finos. Los principales conductores causan un cambio en la población microbiana conduciendo a un incremento en la producción de ALCIGL que inhibe la síntesis de grasa en leche en la glándula mamaria. El resultado es la depresión de grasa en leche. Los principales conductores incluyen muchos de los tipos de grasas erróneas, mucho almidón y bajo pH ruminal. Los sintonizadores finos modifican el contenido de grasa arriba o abajo, pero tienen menos efectos directos sobre la población microbiana del rumen y sobre la síntesis de ALCIGL. Estos incluyen carbonatos K/Na y palmitatos que incrementan el porcentaje de grasa en la leche y levaduras, ionóforos, y alta degradabilidad del almidón que puede disminuir el porcentaje de grasa en la leche. Los sintonizadores finos son utilizados para dar un aumento en la grasa de la leche pero no son un sustituto para la eliminación de los conductores principales que son la raíz de la DGL.
Mucha grasa insaturada
Mucha grasa en la dieta de vacas lecheras es una causa clásica de la DGL. Los nutricionistas están muy conscientes de que la grasa debe ser limitada a menores niveles que la proteína o carbohidratos para evitar el daño a la fermentación del rumen, la reducción del consumo de alimento, y la DGL. Se está intentando empujar el límite en la alimentación de grasa cuando los precios son favorables para los subproductos altos en grasa, cuando los precios de los granos alcanzan niveles record haciendo las grasas comerciales más competitivas o cuando el establo tiene acceso a productos de desperdicio altos en grasa de plantas de procesamiento de alimentos, cercanas. La clave para prevenir la DGL de estos ingredientes altos en grasa es entender completamente el impacto químico y nutricional que estos ingredientes tienen sobre los microbios del rumen y sobre la vaca y escoger una tasa de alimentación que provea el mayor beneficio con el menor riesgo de disminuir la producción de leche y componentes.
Los suplementos de grasa poseen diferentes grados de riesgo de DGL. Las grasas de bajo riesgo son aquellas que causan una pequeña disrupción en la población microbiana del rumen y así mantienen una fermentación normal y una producción limitada de ALCIGL. Las grasas de bajo riesgo generalmente son caracterizadas por un alto contenido de ácidos grasos saturados o por sales de calcio de ácidos grasos. La mayoría de las grasas de sobrepaso comerciales están basadas en estas características, de tal modo que el riesgo de DGL es bajo. La tasa de alimentación de grasa de sobrepaso usualmente es limitada por el costo y disponibilidad. Además, las grasas de sobrepaso son productos secos y sólidos, más que grasas líquidas y son fáciles de empacar, transportar, y mezclar en el establo sin necesidad de equipo especializado. Las grasas de sobrepaso son llamadas grasas inertes en rumen para enfatizar su bajo riesgo de causar disrupción en el rumen.
Los suplementos de grasa de alto riesgo contienen más ácidos grasos insaturados que típicamente son encontrados en forrajes, granos cereales, y semillas oleaginosas (semilla de algodón, soya, canola, girasol, etc.). Los tres principales ácidos grasos insaturados que son consumidos por la vaca son el ácido oleico, linoleico y linolénico con una, dos y tres dobles ligaduras, respectivamente (Figura 5). Una alta concentración de ácidos grasos insaturados en el rumen de una o más de estas fuentes puede inhibir algunas especies microbianas en el rumen. Este cmbio puede favorecer a las especies que producen ALCIGL, la acumulación del cual puede conducir a la DGL. Estos suplementos de grasas insaturadas de alto riesgo son referidos como grasas activas en rumen para enfatizar su tendencia a causar disrupción de las condiciones del rumen.
Una herramienta conveniente para monitorear el consumo de ácidos grasos insaturados riesgosos es llamada RUFAL o Carga de Ácidos Grasos Insaturados en el Rumen. El RUFAL refleja el aporte total de ácidos grasos insaturados en el alimento que entran al rumen cada día. El RUFAL considera los ácidos grasos insaturados de todos los ingredientes del alimento más que solamente los ácidos grasos de los suplementos de grasa. El RUFAL puede indicar mejor el potencial de la disrupción de la fermentación del rumen que simplemente calcular el porcentaje de grasa adicionado a la dieta. Los estudios demuestran que incrementar la RUFAL causa disrupción de la fermentación, la cual puede dañar el comportamiento productivo del animal. La RUFAL excesiva, puede conducir a DGL. Aunque no se ha establecido un nivel de RUFAL exacto que evite la DGL, los valores <3% del CMS se consideran como bajos consumos de grasa, mientras que los valores >3% del CMS indican consumos de ácidos grasos que pueden correr el riesgo de estar muy altos. Hatos con grasa en leche por arriba de 3.8% que tienen un consumo de RUFAL mayor a 3% del CMS, la herramienta solamente sugiere una guía para identificar dietas bajas o altas en grasa.
El pH del rumen
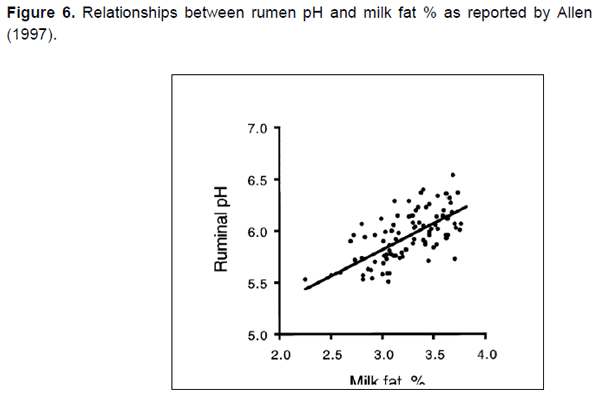
Los estudios muestran que un bajo pH altera la población microbiana en el rumen y causa acumulación de ALCIGL. En un estudio realizado por Fuentes et al. (2009), el pH del rumen disminuyó de 6.5 a 5.5, causando un cambio en la producción de ALC que incluyó un incremento en ALCIGL. Aunque el porcentaje de grasa en leche casi siempre disminuye conforme disminuye el pH del rumen, aún hay mucha variación como dispersión alrededor de la línea en la Figura 6. Esto indica que el pH del rumen no es el único factor controlando el porcentaje de ALCIGL y de grasa en la leche. Así, la acidosis ruminal no debería verse como un pre-requisito para la DGL.
Los factores que pueden resultar en cambios marcados en el pH ruminal en un período de 24 horas incluyen: perfil de carbohidratos dietéticos y las tasas de degradación de las fracciones de los carbohidratos dependiendo de la fuente, procesamiento y humedad; aporte de FDN efectiva físicamente (FDNef), dependiendo de la fuente y el tamaño de partícula; y la producción de buffer en la saliva como una función del aporte de FDNef y la fuente (Shaver, 2005). A pesar de nuestro entendimiento general de estos factores, el grado y la duración de un bajo pH ruminal requerido para causar un flujo suficiente de ácidos grasos insaturados a través de vías alternativas de BH ruminal no se conoce. Aunque los datos son limitados, los cambios en el pH ruminal probablemente son los que más se asocian con la DGL debido a que causan un cambio en la población bacteriana favoreciendo las vías alternativas de BH. El pH del rumen tiene efectos independientes sobre la BH y los intermediarios de BH.
Martin y Jenkins (2002) examinaron las incubaciones de cultivo continuo que fueron conducidas a una tasa de dilución de 0.05 y 0.10/h con valores de pH de 5.5 y 6.5, y 0.5 y 1.0 g/L de carbohidratos solubles mixtos. Ellos encontraron que el factor medio ambiental con mayor influencia sobre BH y perfil de ácidos grasos trans fue el pH del cultivo. En un pH de 5.5, la concentración de trans-C18:1 y ALC fue significativamente reducida resultado de la medida reducida del ácido linoleico BH. Efectos similares fueron observados por Trogeler-Meynadier et al. (2003). Las bajas cantidades de ALC de la reducción en la medida de BH en un pH de 6.0 puede ser debido a baja actividad de isomerasa o a una alta actividad reductasa. Sin embargo, ellos encontraron que un bajo pH (pH 6.0), resultó en bajas cantidades de trans-11 C18:1 en todos los tiempos de incubación comparado con un mayor pH (pH 7.0), pero la concentración de trans-10 C18:1 fue mayor de las 16 a 24 de incubación indicando un cambio en los intermediarios de BH. Un bajo pH inhibió la isomerización inicial y la segunda reducción (trans-11 C18:1 a ácido esteárico), conduciendo a una acumulación de trans-11 C18:1 en cultivos ruminales (Troegeler- Meynadier et al., 2006). Choi et al., (2005) report que cis-9 trans-11 ALC son producidos en un pH mayor a 6.2 por las bacterias del ruen, pero los ALC trans-10 cis-12 son producidos más que los ALC cis-9 trans-11 en un bajo pH. Ellos concluyeron que las bacterias que producen ALC trans-10 cis-12 pueden ser más aero y acido-tolerantes que las bacterias que producen ALC cis-9 trans-11.
Mucho almidón
Mantener adecuados niveles de forraje en las dietas de vacas lecheras disminuye el riesgo de DGL. Como se explicó previamente, el forraje puede ayudar a mantener el pH del rumen y limitar la síntesis de ALCIGL. Este enfoque enfatiza que la FDNef mantiene la masticación y la producción de buffer en la saliva. Los nutricionistas utilizan guías de forrajes específicos para establos específicos con necesidad de forraje individualizado. Dentro de estas referencias, sin embargo, mantener un programa consistente de forraje es la primera línea de defensa contra los problemas de DGL.
Un atributo de los forrajes menos conocido y casi siempre ignorado relacionado a la DGL es su contribución al consumo total de grasa de la vaca. Por ejemplo, los ácidos grasos en el ensilaje de maíz promedian alrededor de 1.5 a 2.0% de la MS. Pero puede alcanzar el 3.5% o mayor. Es importante recordar que el contenido de ácidos grasos no es el mismo que el contenido de grasa cruda cuando solicitamos el análisis del forraje. El contenido de grasa tradicionalmente ha sido determinado como componente del extracto etéreo del alimento. Además, la grasa cruda en granos cereales, forrajes y de la ración total mezclada casi siempre contiene menos del 60% de ácidos grasos. El forraje que contiene 3.5% total de ácidos grasos podría contener de 5 a 6% de grasa cruda.
Dadas las grandes cantidades que se alimentan a las vacas en algunas operaciones, estas aportan consumos significativos de grasa solo del ensilaje de maíz. Los altos consumos de ácidos grasos también se han reportado de forrajes en pastoreo, pero de nuevo, los retos son análisis apropiados. El Ryegrass que pastaron las vacas en la Universidad de Clemson de noviembre a marzo de 2009 tuvo un contenido de ácidos grasos inicial de 6.8% de la MS y cayó a 4.7% al final del pastoreo (Freeman-Pounders et al., 2009). Importante, el análisis del heno no representa consumo de pastoreo. El cortar y secar material de planta durante el proceso de henificación causa una pérdida extensiva de ácidos grasos y otros nutrientes porque el metabolismo de la planta continúa por un tiempo después del corte del zacate. Para representar mejor lo que una vaca consume durante el pastoreo, las muestras de ryegrass en la Universidad de Clemson fueron cortadas e inmediatamente puestas en nitrógeno líquido para detener todo el metabolismo de la planta. Luego, las muestras fueron secadas en frío y se mantuvieron congeladas antes del análisis.
Interacciones entre factores de riesgo
Un cambio repentino en un parámetro nutricional, aún dentro de los requerimientos según las guías, pueden desbalancear el medio ambiente ruminal completo y causar la acumulación de ALCIGL. Así, si estás dentro de los requerimientos, pero aun así hay DGL, entonces el parámetro global de todos los parámetros ha sido dañado.
Por ejemplo, los cultivos de microorganismos del rumen fueron alimentados a dietas altas en maíz o cebada con la presencia o ausencia de aceite de soya (0 y 5%) y la presencia o ausencia de monensina (0 y 25 ppm). La adición de aceite de soya incrementó las concentraciones de trans-10 18:1 en los cultivos de las dietas maíz y de cebada (Jenkins et al., 2003). En menor grado, la monensina también incrementó las concentraciones de trans-10 18:1 en las dietas de maíz y de cebada. Sin embargo, cuando la monensina y el aceite de soya se adicionaron juntos a la dieta, la combinación interactuó. La adición de monensina con aceite de soya no elevó el trans-10 18:1 en dietas basadas en maíz. En dietas basadas en cebada, la adición de monensina con aceite de soya elevó el trans-10 18:1 más que cualquiera de los riesgos por separado.
La interacción de un grano similar, monensina y grasa fue examinada en vacas lecheras lactantes (Van Amburgh et al., 2008). Ochenta vacas Holstein fueron asignadas a una dieta alta (27.7%) o baja (20.3%) en almidón por 21 días, seguido de la adición de monensina (13 ppm) o aceite de maíz (1.25%) por otros 21 días adicionales. Después, se les cambió la dieta a las vacas con niveles opuestos de aceite de maíz por un periodo final de 21 días, con lo que fueron ocho tratamientos. El nivel de aceite fue un factor mayor de riesgo para la DGL comparado con la monensina: el aceite de maíz disminuyó la grasa en leche de 3.32 a 2.99% contra 3.20 a 3.11% por la monensina. La alimentación de dietas altas en almidón tiene efectos limitantes sobre la DGL: la grasa en leche disminuyó de 3.25 a 3.06%. la degradabilidad del almidón pudo haber contribuido a la depresión de DGL en este estudio porque las dietas contenían maíz rolado, el cual tiene una rápida tasa de degradación del almidón en el rumen. Así la degradabilidad, compuesta por el alto consumo de materia seca, puede ser un factor de riesgo más potente sobre la DGL que el sólo consumo de almidón.
SINTONIZADORES FINOS QUE INCREMENTAN LA GRASA EN LA LECHE
Carbonato de Potasio
En 2012 fue completado un estudio que evaluó la relación del nivel de K alimentado en lactación temprana, incrementando DCAD con carbonato de potasio sesquihidratado (Harrison et al., 2012). Las vacas estuvieron en el estudio de 15 a 85 DEL. Las dietas fueron formuladas para tener el mismo nivel de nutrientes, excepto por el K con niveles de K de 1.3% y 2.1% de MS; y niveles de DCAD de 25 y 42 meq/100g MS. La inclusión de una alta cantidad de K en la lactación temprana resultó en un incremento en la producción de leche, leche corregida a 3.5% de grasa y grasa en leche. Este incremento no estuvo asociado con un incremento de consumo de materia seca. Y además parece no estar relacionado al consume de energía.
Las muestras de leche de la mitad de las vacas en cada tratamiento que representó un rango de baja a alta producción fue seleccionada para la caracterización de ácidos grasos en leche. El carbonato de potasio adicionado a la dieta disminuyó los ácidos grasos insaturados y trans, e incrementó el C18:0 en leche. Un estudio reciente de Alonso-Avila et al. (2017) no se encontró efecto significativo de la suplementación de carbonato de potasio sobre el porcentaje de grasa o de trans-10 C18:1 en vacas en lactación temprana que fueron alimentadas con 2% de aceite de soya. Es importante resaltar que la diferencia numérica en el estudio de Alonso- Avila et al. (2017) mostró que el carbonato de potasio incrementó la grasa en leche de 3.04 a 3.42% y redujo el ácido trans-10 C18:1 en leche de 1.34 a 0.56% del total de ácidos grasos.
Una explicación para el incremento del porcentaje de grasa en la leche con el incremento de carbonato de potasio puede estar ligado al pH del rumen. El incremento de K y el pH en el rumen probablemente altera los tipos y la cantidad de intermediarios de la BH producidos por la población microbiana del rumen, la cual a su vez incrementa la grasa en la leche. Un estudio en cultivo continuo realizado por Jenkins et al. (2014) mostró que la adición de carbonato de potasio redujo la producción diaria de trans-10 C18:1 y ALC trans-10, cis-12.
Grasa alto palmítico
La adición de suplementos de grasa con alto contenido de ácido palmítico ha incrementado el porcentaje de grasa en leche en un número de estudios. Los ejemplos incluyen incrementos en la grasa de la leche de 3.44 a 3.93% por la alimentación de 412 g/d de C16 (Mosley et al., 2012), el incremento de grasa en la leche de 3.88 a 4.16% por la alimentación de 361 g/d de C16 (Lock et al., 2013), y un incremento en la grasa de la leche de 3.29 a 3.40% por la alimentación de 545 g/d de C16 (Piantoni et al., 2013. En algunos casos, grasas con menor porcentaje de C16 también incrementó la grasa en la leche. Por ejemplo, el porcentaje de grasa en leche incrementó de 3.37 a 3.67% cuando se alimentó 1.5% de suplemento de grasa con un contenido de 54% de C16 y 34% de C18 (Wang et al., 2010). No se encontró efecto sobre la producción de grasa en la leche cuando se alimentó C18 (Boerman et al., 2017). En algunos casos, los suplementos altos en C16 no tuvieron efecto sobre la producción de grasa en leche (Rico et al., 2014).
Un reporte reciente (de Souza y Lock, 2017) de alimentación de 1.5% de un suplemento alto en C16 a vacas lactantes frescas (0 a 24 d) o en el pico de producción (25 a 67 d) se presentó en la reunión anual del ADSA 2017 en Pittsburgh, PA. Para las vacas frescas, el suplemento de C16 no tuvo efecto sobre la producción de leche, incrementó la grasa en leche en 0.41 unidades porcentuales, pero causó una pérdida de la condición corporal que promedió 41 kg. Para las vacas en el pico de producción, el suplemento de C16 incrementó la producción de leche en 3.4 kg/d, la grasa en leche en 0.21 unidades porcentuales, pero no tuvo efecto sobre la pérdida o ganancia de condición corporal. El resultado podría sugerir que la alimentación de alto C16 podría tener una partición de energía diferente dependiendo del estado de lactación.
SINTONIZADORES FINOS QUE DISMINUYEN LA GRASA EN LA LECHE
Levaduras/hongos
Las levaduras y los hongos se consideran factores significativos de riesgo para DGL, pero se sabe poco exactamente acerca de cómo afecta la función del rumen y la acumulación de ALCIGL. Las teorías acerca de los hongos y levaduras sugieren que pueden producir sustancias antimicrobianas como parte de su metabolismo, el cual a su vez puede impactar de manera negativa la población microbiana del rumen; sin embargo, mucho permanece sin probarse. Altos conteos de hongos y levaduras en alimentos fermentados son indeseables, no solo por el riesgo de la DGL, sino también porque puede reducir el consumo de alimento, afecta negativamente la salud animal, y disminuye el comportamiento productivo general, además de que causa pérdidas adicionales en el alimento a través del desperdicio de éste. En ensilajes bien preservados, el conteo de levaduras es menor a 10,000 UFC/g. los conteos que afectan la salud animal y el comportamiento productivo no están bien definidos y probablemente dependen de cepas específicas de hongos o levaduras que infectan la planta. Como una regla general, los conteos de levadura por arriba de un millón de UFC/g deberían causar preocupación.
Altas tasas de degradación del almidón
La tasa de degradación del almidón en los granos también determina el riesgo de DGL. Observaciones de campo e inferencias de estudios indican que la tasa de fermentación rápida del almidón está ligada a un mayor riesgo de DGL. Cuando se utilizó maíz procesado con más del 80% de fermentabilidad en una prueba in vitro de 7 horas, la producción diaria de ALCIGL incrementó comparado a la adición de maíz no procesado con el 48% de fermentabilidad del almidón (Lascano et al., 2016). Ellos reportaron cambios en los intermediarios de la BH (expresada como porcentaje del total de isómeros) de baja a alta Kd que incluyó reducciones en ALC trans-11 18:1 y cis-9, trans-11 pero incrementó en ALC trans-10 18:1 y trans-10, cis- 12. Los resultados mostraron que el incremento en la Kd del almidón en cultivo continuo mientras se mantiene un nivel constante de almidón causa una elevación de los intermediarios de BH ligados a la DGL.
Los ingredientes fermentados con alto contenido de grano tal como el ensilaje de maíz y el maíz alta humedad tienen el mayor riesgo. Las diferencias en las variedades de maíz, el tiempo de almacenamiento en el silo, y las condiciones climáticas para el crecimiento de las plantas pueden conducir a rápidas tasas de degradación del almidón en el ensilaje y el maíz alta humedad. El mayor tiempo de almacenamiento puede conducir a altas tasas de degradación del almidón. Un estudio desarrollado por Newbold et al. (2006), utilizando una prueba in vitro en fluido de rumen durante 3 horas, encontró un 30% de incremento en la degradación del ensilaje de maíz almacenado por 2 meses contra 10 meses. Si se sospecha de altas tasas de degradación de almidón como una causa de la DGL, usualmente hay poco que se pueda hacer para remediar la situación. Una opción es diluir el forraje con un alimento menos degradable, pero es poco probable que esté disponible. Una opción es enfocarse en otros factores de riesgo (tal como pH del rumen y grasa dietética) para minimizar la producción de ALCIGL.
Monensina
Los ionóforos disrumpen la BH ruminal de manera similar a los suplementos de grasa insaturada. La mayor concentración de ácido linoleico, trans C18:1 y ALC se mantuvo en una prueba de cultivo continuo de bacterias ruminales siguiendo la infusión de monensina, nigericina o tetronasina (Fellner et al., 1997). Alimentar monensina tuvo efectos similares al suplemento de grasa sobre el incremento del ácido linoleico y AG trans en leche de vacas lactantes, y también causó una reducción en el porcentaje de grasa en la leche (Sauer et al., 1998).
Debido a la naturaleza de las micotoxinas y otros ionóforos, es razonable esperar que funcionen de acuerdo a la teoría de la BH y causar un cambio microbiano en el rumen que altera las vías de la BH. La monensina incrementa los requerimientos de mantenimiento de las bacterias gram positivas en el rumen lo cual hace a esta bacteria menos competitiva en el ambiente ruminal (Duffield y Bagg, 2000). Los microorganismos ruminales capaces de la hidrogenación de ácidos grasos son casi siempre divididos en Grupos A y B basados en sus productos finales y patrones de isomerización durante la BH (Harfoot y Hazlewood, 1997). Las especies de bacterias en el grupo A hidrogenan ácido linoleico a trans 18:1 pero parece ser incapaz de hidrogenar monenes. Las bacterias del grupo B pueden hidrogenar un amplio rango de monenes, incluyendo trans-11 C18:1, a ácido esteárico. El pequeño grupo de bacterias en el grupo B son gram positivas.
Así, disminuyendo el número de bacterias que pueden llevar a cabo este proceso potencialmente conducen a la generación de intermediarios de la BH en el rumen. Esto fue descubierto por Fellner et al. (1997) cuando examinaron el efecto de la monensina siguiendo la infusión continua de ácido linolénico en fermentadores ruminales. Cun una dieta suplementada la tasa de formación de 18:0 fue de 7.5 mg/L/h mientras que disminuyó a solo 2.7 mg/L/h cuando la monensina fue suplementada (Fellner et al. (1997). Es importante recorder, sin embargo, que un increment en el flujo ruminal de intermediaries de la BH no será un problema si la vía típica de la BH está presente. Sin embargo, aun si una pequeña proporción de PUFA dietético está siendo hidrogenada a través de la vía que produce ALC trans- 10, cis-12 e intermediarios relacionados, la monensina puede potencialmente incrementar el riesgo de DGL.
Conclusiones
El conocimiento de la DGL ocurrió con el descubrimiento de que ésta está relacionada con la producción de ALC en el rumen. Utilizar controles de manejo de la DGL limitando la acumulación de ALCIGL en el rumen. En general, no hay un único factor dietético que sea responsable por la DGL, y las interacciones entre varios componentes dietéticos puede incrementar el flujo ruminal de ALCIGL. Todos los riesgos tienen que ser considerados en relación a la combinación de factores que juegan una formulación de una ración determinada y con relación a las limitaciones de manejo y de la planta física. Investigaciones futuras son requeridas para un mejor entendimiento de las condiciones del rumen que promueven la formación de ALCIGL que puede disparar la DGL. Un entendimiento mejorado de estos eventos proveerá el marco crítico con el cual se resolverán mejor los problemas de DGL.