Introducción
Los carbohidratos (CHO) son los principales constituyentes de la materia seca (MS) de los forrajes, entre los que predomina la celulosa, seguida de hemicelulosa. En consecuencia, la actividad celulolítica es fundamental para la nutrición de los rumiantes y herbívoros altamente dependientes del forraje. Sin embargo, la fermentación amilolítica cobra importancia creciente con el consumo de CHO no estructurales, ya sea como constituyentes del forraje o suministrados vía suplementos.
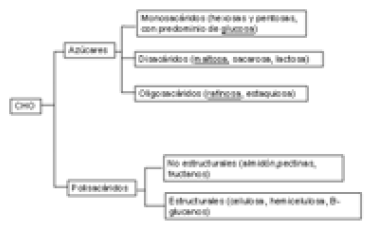
Clasificación y contenido de CHO en alimentos.
Los principales CHO constituyentes de los alimentos del rumiante se describen en la Figura 1.
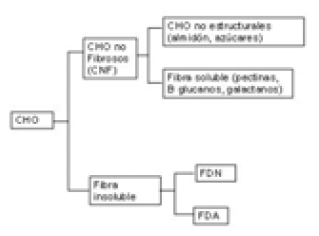
Las fracciones analíticas en que se insertan los diferentes tipos de CHO se presentan en la Figura 2.
Figura1. Principales carbohidratos constituyentes de los alimentos.
Figura 2. Fracciones analíticas de los carbohidratos
El grupo de los azúcares está constituido por tres categorías:
Monosacáridos:En los CHO solubles de los pastos están representados por fructosa y glucosa, en una relación aproximada de 2:1, y constituyen sustratos para la síntesis de sacarosa y oligosacáridos.
Disacáridos: Principalmente constituidos por sacarosa, la que en los pastos representa alrededor de 15% de los CHO solubles, siendo también utilizada por la planta en la formación de oligosacáridos. La sacarosa puede ser alta al suministrar melaza. La maltosa se produce al hidrolizar almidón.
Oligosacáridos: Son derivados de la sacarosa y pueden tener de 3-9 unidades de monosacáridos, principalmente galactosa y fructosa. Son los principales componentes de los CHO solubles, alcanzando en ballicas el 60% de esta fracción. También son CHO de reserva en semillas de leguminosas. Fermentan con facilidad, pero no pueden ser digeridos por carbohidrasas intestinales.
Los CHO solubles pueden superar el 20% de la MS en gramíneas, especialmente en variedades de ballicas ricas en azúcares, pudiendo ser responsables de cuadros de acidosis en condiciones de pastoreo.
En el grupo de los polisacáridos destacan los siguientes:
Almidón:Es el principal CHO de reserva en plantas superiores, depositado en forma de gránulos en semillas, raíces y tubérculos. Los gránulos son relativamente densos, insolubles y se hidratan poco en agua fría. Está compuesto por dos polímeros, amilosa (lineal) y amilopectina (ramificado), que constituyen el 25 y 75%, respectivamente, en la mayoría de los almidones. La digestibilidad del almidón aumenta con el contenido de amilopectina.
Celulosa: Está compuesta por cadenas lineales de glucosa (enlaces B 1,4), por lo cual es solo degradable por fermentación. Su apariencia fibrilar en la pared celular se debe a la agrupación paralela de cadenas de celulosa unidas por puentes de H, lo que determina una estructura cristalina difícil de hidratar y muy insoluble. Los B-glucanos, son cadenas de glucosa igual que la celulosa, pero son fermentados rápidamente. Se encuentran en gramíneas y en la fibra de granos de cereales.
Hemicelulosa: Es un polisacárido complejo,química y estructuralmente distinto de la celulosa, constituido por hexosas y pentosas (enlaces B 1,4) y contribuye al fortalecimiento y flexibilidad de la pared celular. Se encuentra formando enlaces cruzados con celulosa, lignina y proteína en la pared celular. Solo es degradable por enzimas de microorganismos.
Pectinas:Son CHO coloidales ramificados de la pared celular, de alto peso molecular, también consideradas como “cemento intercelular”. El principal componente es ácido d-galacturónico, esterificado en distintas proporciones con metanol y diferentes azúcares unidas a cadenas laterales. Son solubilizadas por el Detergente Neutro y no aparecen en el análisis de fibra insoluble, por lo que se rescatan en la fracción soluble. Su contenido es mayor en plantas leguminosas que en gramíneas y es abundante en frutos blandos y pared celular de raíces como la remolacha. Son degradables por enzimas de microorganismos, pero fermentan rápidamente.
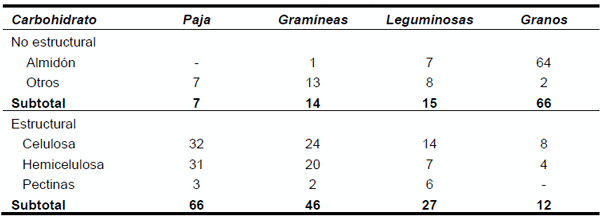
La distribución de los diferentes CHO en forrajes y granos se presenta en el Cuadro 1.
Cuadro 1. Contenido de carbohidratos en alimentos (% BMS).
Los forrajes toscos, poseen principalmente celulosa y hemicelulosa; las gramíneas en el follaje contienen más azúcares, más celulosa y hemicelulosa y menos pectinas. En el follaje de leguminosas hay más almidón que en el de gramíneas. Los granos de cereales contienen principalmente almidón.
Existen diferencias importantes en el contenido y distribución de monosacáridos entre alimentos. En henos y pajas, sobre el 90% de la glucosa se encuentra el la pared celular, en cambio, en ensilaje de maíz solo el 54% y la diferencia (46%) se encuentra en la fracción soluble. El resto de los monosacáridos, exceptuando xilosa y arabinosa (componentes de hemicelulosa), se encuentra en la fracción soluble, presumiblemente como componentes de las pectinas.
Carbohidratos y la función ruminal
Los alimentos poseen ciertas propiedades, resultantes de la cantidad y tipo de CHO que los componen. Entre estas se incluye la capacidad de retener agua, iones y otras sustancias solubles, atribuible principalmente a grupos activos de los diferentes CHO y a aspectos de su estructura. Estos grupos facilitan la interacción con el agua, iones inorgánicos, proteínas y lípidos y que se traducen en los efectos relacionados con la adsorción (agua y otras sustancias), intercambio iónico y capacidad tampón. La capacidad tampón o buffer de los alimentos depende principalmente del contenido de fibra y de la capacidad de intercambio de los componentes de la misma, y en menor grado de componentes de la fracción soluble, como la proteína. Del Cuadro 2, que resume los grupos activos en carbohidratos, proteínas y lignina y su capacidad de acción en la fermentación ruminal, se desprende lo siguiente:
Celulosa. Posee grupos alcohólicos (OH) muy involucrados en puentes de hidrógeno (H+), lo que restringe su capacidad de retención de hidratación. No posee capacidad de intercambio ni efecto tampón.
Almidón. Debido a sus grupos alcohólicos, no posee actividad de intercambio ni capacidad tampón; si bien se hidrata más que la celulosa, posee una baja hidratación en agua fría.
Hemicelulosa. Posee capacidad de retención de agua aunque su capacidad tampón y de intercambio es inferior que la de pectinas y lignina.
Pectinas. Poseen una alta capacidad de hidratación y además una significativa capacidad tampón y de intercambio iónico, siendo efectivas en el control del pH ruminal. También participan en la adsorción de lípidos, lo que posibilita la eliminación de ácidos biliares y colesterol vía fecal.
Proteínas. La capacidad de hidratación dependerá de la disponibilidad de grupos carboxilos libres. Pueden fijar grasas y ácidos biliares, lo que se favorece por el contenido de Nitrógeno que permite el intercambio aniónico.
Lignina. A pesar de su nula contribución nutricional, posee una importante capacidad de intercambio, efecto tampón, de hidratación y de adsorción de lípidos.
Cuadro 2. Grupos activos y propiedades funcionales de carbohidratos, lignina y proteínas en el rumen.
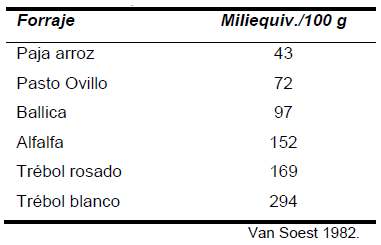
La capacidad de hidratación, de intercambio y el efecto tampón es baja en forrajes altamente celulósicos como la paja y en gramíneas en madurez avanzada. Comparativamente, la capacidad de intercambio es más baja en gramíneas que en leguminosas, lo que se explica en gran medida porque las leguminosas poseen una fibra más lignificada, con menos celulosa y más pectinas, además de un mayor contenido medio de proteína (Cuadro 3).
De lo anterior se deduce que la combinación de forrajes celulósicos con almidón no favorece el desarrollo de buenas condiciones de funcionalidad ruminal, por una baja capacidad de hidratación y de intercambio iónico y baja actividad tampón, siendo relevante el efecto positivo que ejercen las leguminosas y alimentos ricos en fibra soluble, en crear un favorable ambiente ruminal.
Cuadro 3. Capacidad de intercambio de diferentes forrajes.
Carbohidratos y la fermentación ruminal
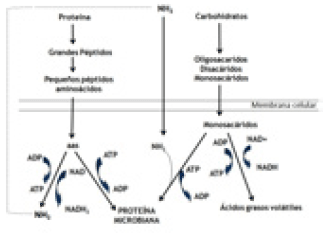
La fermentación ruminal de CHO y de proteínas es interdependiente ya que los monosacáridos liberados por las enzimas microbianas, proveen de energía (ATP) para el metabolismo de los microorganismos (m.o.), los cuales, con este aporte más la proteína del alimento se reproducen y aumenta su biomasa o la producción de proteína microbiana (Figura 3).
Figura 3. Relación entre el metabolismo de los carbohidratos y de las proteínas en el rumen.
Etapas y rutas metabólicas.
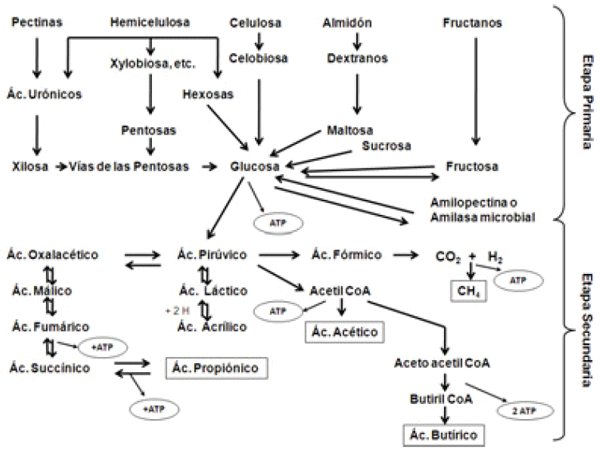
En la fermentación, se distinguen dos etapas (Figura 4):
Etapa primaria. Consiste en la liberación de los componentes unitarios de los macronutrientes (predominantemente glucosa a partir de CHO y aminoácidos y amonio a partir de proteínas y NNP), por enzimas microbianas extracelulares de especies productoras.
Etapa secundaria. Absorción por los m.o. (especies utilizadoras) de los componentes unitarios liberados y metabolismo intracelular de estos, proceso que es funcional a su nutrición y proliferación. El metabolismo microbiano además recicla factores esenciales al grupo anterior y posibilita la transferencia de H+entre especies productoras y utilizadoras. Las rutas metabólicas de los microorganismos son similares a las empleadas en el metabolismo del animal. El intermediario clave, que conecta las diferentes rutas metabólicas es el piruvato, formado de glucosa el que ingresa rápidamente a las bacterias.
En condiciones anaeróbicas, los microorganismos no pueden oxidar los sustratos hasta CO2 y agua, por lo que queda una proporción importante de estos retenida en los productos finales de la fermentación (AGVs y proteína microbiana), lo cual favorece la nutrición del animal. Sin embargo, con la dieta varían el pH y la concentración y proporciones relativas de diferentes microorganismos que participan, y también la generación de productos finales de la fermentación. La fermentación de aminoácidos también produce AGVs (isoácidos ramificados) que son usados para formar nuevos aminoácidos. Los gases son eliminados por eructación lo que representa una pérdida de energía. El calor producido también se pierde, a menos que se requiera para mantener la temperatura corporal en ambiente frío.
Con forrajes el pH ruminal es más elevado (6,2 - 6,8) y predominan m.o. celulolíticos; al bajar el pH gradualmente aumentan los amilolíticos, para finalmente predominar los lactobacilos cuando el pH baja de 5,5.
Figura 4. Etapas y rutas metabólicas en la fermentación de carbohidratos (Mod.Grudsky y Arias 1983.)
Formación de acetato. El acetato es el producto predominante en el balance de AGVs, debido a que en la fermentación anaeróbica, hay necesidad de reacciones que permitan utilizar el exceso de H+ que se produce, el cual es transferido a especies utilizadoras de H+, en que destacan las metanogénicas. La producción de metano estimula una mayor producción de acetato y CO2 e H+y por ende, la continuidad de la fermentación. Ello explica que exista una reducción drástica de productos reducidos como ácido láctico, succinato, propionato, etanol y formato, en favor de acetato y CO2, y el predominio de acetato en el balance de AGVs. La producción de acetato es favorecida por alimentación rica en fibra, con participación de diferentes especies de bacterias celulolíticas.
Formación de propionato. La producción de propionato ocurre con participación de diferentes especies de bacterias amilolíticas. También hay bacterias celulolíticas que poseen actividad amilolítica.
Con alimentación rica en forrajes (baja ingesta de CHO no estructurales), la producción de propionato es reducida y no está asociada a producción de lactato como producto intermediario. En este caso, la principal ruta metabólica es vía succinato. Una segunda ruta alternativa (lactato-acrilato), que es favorecida por dietas ricas en concentrados, o azúcares, implica un aumento en producción de propionato. Esta ruta puede aumentar el ácido láctico ruminal, que podrá ser convertido en propionato con pH ruminal >6,2. De lo contrario, el lactato se acumula generando un cuadro de acidosis. Con dietas ricas en forraje, la primera ruta explica más de 90% del propionato formado. En cambio, con dietas ricas en concentrado, la segunda ruta aumenta su contribución a más de 20%.
Relación acetato:propionato (A/P): Existe una relación inversa entre la producción de acetato y de propionato, aunque difícilmente la producción de propionato llegará a superar la producción de acetato. Al aumentar los forrajes, 70% o más del total de AGVs corresponde a ácido acético, proporción que se reduce al disminuir el forraje y aumentar los concentrados. La misma tendencia ocurre al aumentar la calidad (digestibilidad) de los forrajes.
Formación Ácido láctico y alcoholes: El lactato a nivel ruminal puede tener los siguientes origines:
a) Vía metabólica alternativa (lactato-acrilato) con dietas ricas en azúcares y/o almidón
b) A partir de propionato, transformado en la pared ruminal (aprox 30% del total)
c) A partir de la ingesta, principalmente de ensilajes (hasta 0,8 kg/d en vacas con ensilaje de buena calidad fermentativa, ad-libitum).
El ácido láctico es metabolizado principalmente a propionato, acetato y CO2 y también ácido butírico. La acumulación observada al suministrar dietas ricas en almidón o azúcares, demuestra que el ácido láctico no es un importante intermediario y que los microorganismos no poseen una gran capacidad de metabolizarlo.
El etanol ruminal debe provenir principalmente de la ingesta de alimentos que lo contienen debido a que la producción de alcohol no es favorecida en la fermentación ruminal, por requerir ATP. Importantes fuentes de alcohol pueden ser ensilajes de maíz (hasta 3% BMS), ensilaje de grano húmedo (hasta 2% BMS) y ensilajes directos de forrajes en general (hasta 1,0% BMS), cuando las condiciones favorecen el desarrollo de levaduras. Las levaduras también degradan ácido láctico cuando el silo y queda expuesto al aire. La ingesta diaria de alcohol con ensilaje ad-libitum puede superar 0,2 kg/d en vacas lecheras.
El alcohol es poco usado por las bacterias ruminales, pudiendo contribuir a cierta formación de acetato, siendo una fuente menor de energía. La mayor parte del alcohol es absorbido y metabolizado por el animal de igual modo que en no rumiantes.
Eficiencia ruminal versus eficiencia del animal
La cantidad de CHO convertida anaeróbicamente en AGVs, es mucho mayor que la convertida en microorganismos. Comparativamente, la producción de AGVs rinde a la fermentación mucho menos ATP que su oxidación en los tejidos del animal.
Las sustancias que permiten captar H+ y generar menos pérdidas de C y CO2(ácidos láctico, propiónico, fórmico) favorecen la eficiencia para el animal. Sin embargo, dietas que deprimen la producción de acetato a favor de propionato (vía lactato), reducen la disponibilidad de energía (ATP) para la fermentación y se resiente la producción de proteína microbiana. En consecuencia, al estrecharse la relación A/P aumenta la disponibilidad de energía para el animal y se reduce la disponibilidad para la fermentación.
Según el Cuadro 4, con raciones concentradas (relación A/P 1:1), la disponibilidad de energía para la fermentación (8 ATP/mol glu) es un 33% inferior que con una dieta rica en forraje (12 ATP/mol glu), lo que demuestra la conveniencia de suministrar dietas ricas en fibra digestible para favorecer la disponibilidad de ATP en el rumen.
Destino de los productos de la fermentación de CHO
La mayor parte de los AGVs se absorbe en el rumen (hasta 4 kg/día) y en menor grado en retículo y omaso, pudiendo cierta cantidad pasar al abomaso y ser absorbida a nivel intestinal. La producción de gases puede ser muy rápida (hasta 30 L/hora) en una mezcla constituida aproximadamente por 40% CO2, 30-40% CH4 y 5% H+, lo que explica el riesgo de meteorismo (timpanismo) agudo con dietas ricas en concentrados.
En general, la velocidad de absorción aumenta con el largo de cadena, por lo que sigue el siguiente orden: butírico>propiónico>acético. La absorción es más rápida cuando baja el pH ruminal, debido a que los AGVs están en forma no disociada (difusión pasiva).
Cuadro 4. Disponibilidad de energía según producto la fermentación.
El ácido butírico es metabolizado en la pared ruminal y en el hígado a βHB; aproximadamente 30% del ácido propiónico es transformado en la pared ruminal a ácido láctico y el resto es transportado como tal al hígado para ser metabolizado a glucosa; la mayor parte del acetato se absorbe como tal y es usado por los tejidos y glándula mamaria; el ácido láctico se absorbe lentamente, a solo un 10% de la tasa de absorción de los AGVs y es conducido al hígado para ser metabolizado a glucosa.
Fuentes de CHO no-fibrosos y características
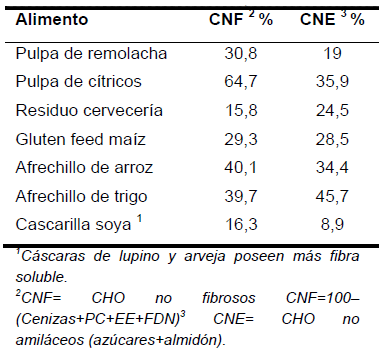
Algunas características ligadas al uso de fuentes de CNF (granos de cereales, subproductos ricos en fibra soluble, se presentan en los Cuadros 5, 6 y 7).
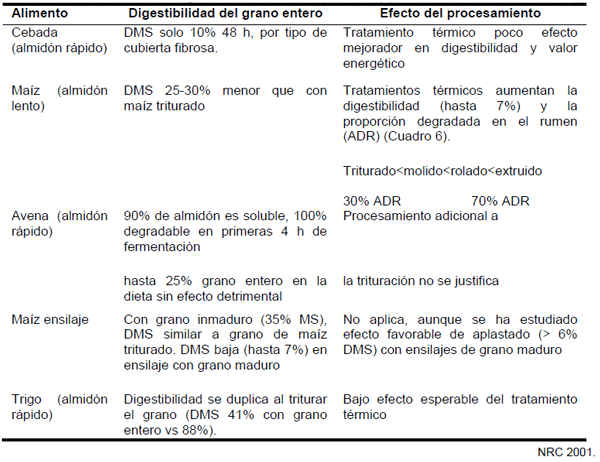
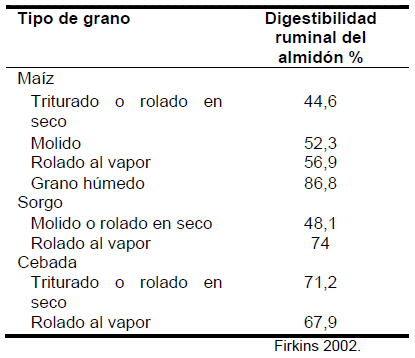
Al aumentar el consumo de las vacas, la tasa de pasaje de nutrientes al intestino aumenta. Investigaciones demuestran que 5-20% del almidón de maíz consumido se digiere después del rumen, principalmente en el intestino delgado, lo que puede representar más de 4 kg/día. Ello dependerá esencialmente del tipo de grano y del procesamiento del mismo, principalmente cuando es con calor húmedo (Cuadros 5 y 6).
Alimentos ricos de fibra soluble (Cuadro 7), son atractivos para la nutrición ruminal debido a que entregan energía en forma rápida, con ácidos débiles (predomina acético), ejercen efecto tampón y por lo tanto su uso genera bajo riesgo de acidosis. Sin embargo, estos alimentos contienen alta cantidad de FDN rápidamente fermentable, pero niveles marginales de fibra efectiva, por ello se limitan a no más de 30-35% en la dieta.
La fibra soluble en el Cuadro 7, está conformada por los CHO no fibrosos, más gran parte de los azúcares de la fracción de CNE, que corresponden a oligosacáridos del tipo B glucanos y galactanos.
Cuadro 5. Características de granos de cereales sin procesar y procesados.
Cuadro 6. Proporciones de almidón degradado en el rumen e intestino según forma de procesamiento.
Cuadro 7. Contenido de CHO no fibrosos y fibra soluble de diferentes subproductos.