Factores que influyen en el Valor Nutricional de las Grasas utilizadas en las Dietas para Bovinos de Engorda en Confinamiento: Una Revisión
Publicado: 29 de diciembre de 2006
Por: Alejandro Plascencia Jorquera, Germán D. Mendoza Martínez, Carlos Vásquez Peláez y Richard Avery Zinn
Excluyendo las interacciones asociativas, el valor de EN de las grasas alimenticias es una función altamente relacionada con su digestibilidad intestina. Por lo anterior, una gran atención se ha dirigido para comprender los factores intrínsecos (naturaleza y calidad de la grasa) y extrínsecos (método de adición y nivel de consumo) que afectan su valor como alimento animal. De esos factores, el nivel de consumo de grasa ha demostrado ser el de mayor impacto. Cuando el consumo total de grasa supera la proporción de 0.96 g de grasa/kg de peso vivo, el valor energético de la grasa disminuye en forma lineal como resultado directo de la disminución de la digestibilidad intestinal de los ácidos grasos, principalmente C18:0; lo anterior, posiblemente resultado de una limitada capacidad de producción biliar en el bovino cuando consume altas cantidades de lípidos.
Summary
Summary
Except for associative interactions, the NE value of dietary fat is largely a function of its intestinal digestibility. Thus, attention has been directed to understanding intrinsic (nature and fat quality) and extrinsic (fat method addition and inclusion level) factors that influence its value as feed. Among these factors, fat intake level has the greatest impact on its energy value. When total fat intake exceeds 0.96g/kg BW, the NE of fat decreases linearly, as a result of the decline in postruminal fatty acid digestibility, mainly of C18:0; the latter probably explained by a limited capacity of bile production in the ruminant at high levels of fat intake.
Resumo
Excluindo as interações associativas, o valor de EN das gorduras alimentícias é uma função altamente relacionada com sua digestibilidade intestinal. Pelo anterior, uma grande atenção se tem dirigido para compreender os fatores intrínsecos (natureza e qualidade da gordura) e extrínsecos (método de adição e nível de consumo) que afetam seu valor como alimento animal. Desses fatores, o nível de consumo de gordura tem demonstrado ser o de maior impacto. Quando o consumo total de gordura supera a proporção de 0.96 g de gordura/kg de peso vivo, o valor energético da gordura diminui em forma linear como resultado direto da diminuição da digestibilidade intestinal dos ácidos graxos, principalmente C18:0; o anterior, possivelmente resultado de uma limitada capacidade de produção biliar no bovino quando consome altas quantidades de lipídeos.
Las grasas y aceites son una fuente alimenticia para rumiantes, de alta densidad energética y de bajo costo. Los valores de contenido energético (EN) para bovinos de engorda asignados por los estándares actuales (NRC, 1996) son de 6,00 y 4,75Mcal·kg-1 para mantenimiento y ganancia, respectivamente. Las grasas de grado alimenticio contienen aproximadamente 90% de ácidos grasos totales (AFOA, 1999) y éstos representan casi el 100% de su contenido energético (Zinn, 1989a). Por ello el valor energético de las grasas está supeditado a la digestibilidad de sus ácidos grasos, la cual se ha determinado en 80% en especies rumiantes (Palmquist, 1991). Aún así, los resultados generados en diversos estudios muestran a la grasa alimenticia como uno de los insumos de mayor variabilidad en cuanto a su valor nutricional, por lo que una gran atención se ha dirigido a comprender los factores que lo afectan. Las áreas de énfasis incluyen tipo o fuente de grasa (Zinn, 1989b; Brandt y Anderson, 1990; Khrebiel et al., 1995), contenido de ácidos grasos libres (Zinn, 1992; Zinn et al., 2000), grado de saturación (Johnson y McClure, 1972; Elliott et al., 1997; Plascencia et al., 2001), método de adición (Zinn et al., 1998; Zinn y Plascencia, 2004a) y nivel de adición (Haaland et al., 1981; Moore et al., 1986; Zinn 1989a, 1994; Pylot et al., 2000; Plascencia et al., 2002).
El objetivo de esta revisión es discutir los principales factores que influyen en el valor nutricional de las grasas utilizadas en las dietas para bovinos de engorda en confinamiento. Éstos incluyen la fuente de grasa, así como el método y nivel de adición.
Fuentes y Tipos de Grasas
Fuentes y Tipos de Grasas
Los lípidos de los forrajes se encuentran principalmente en forma de ácidos grasos poliinsaturados esterificados como galactosilglicéridos. La concentración de ácidos grasos en esta forma rara vez supera el 1,5% de la materia seca de la dieta. En cambio, los ácidos grasos contenidos en cereales, semillas oleaginosas y grasas libres es variable, más elevado y en forma de triglicéridos.
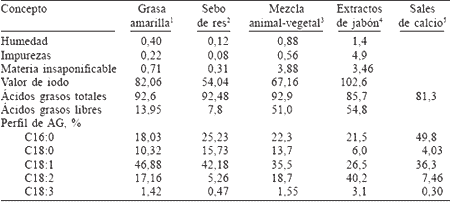
En relación a las grasas libres, son diversas las fuentes de grasa que son utilizadas en la alimentación de bovinos de engorda. Éstas difieren principalmente en contenido de impurezas, de ácidos grasos libres (AGL) y en el grado de saturación (Tabla I). A continuación se describen brevemente las fuentes de grasas más comúnmente utilizadas.
Grasa amarilla
El término de "amarilla" se debe a su apariencia. También se le conoce como grasa de restaurante o grasa de cocina, ya que su origen es de cualquier combinación de los desperdicios o sobrantes de grasas y aceites colectados en cafeterías, restaurantes de comida rápida y panaderías. Como resultado de cocinar cada vez más con aceites vegetales, la mayor parte de grasa amarilla recobrada es de origen vegetal que ha sido parcialmente hidrogenizada para un mejor desempeño en el proceso de cocinado, de tal forma que la proporción de insaturados:saturados es de 2,6 aproximadamente (Zinn, 1988; Plascencia et al., 1991). Debido a la diversidad de sus fuentes, la grasa amarilla no es muy uniforme en su composición y puede variar de un área a otra, o de una planta a otra. De acuerdo a los parámetros establecidos por la Asociación Americana de Grasas y Aceites (AFOA, 1999), su punto de fusión debe ser menor a 40ºC y no debe contener más de 15% de AGL y un máximo de 2% de impurezas.
Sebo
El sebo o grasa animal es un subproducto derivado principalmente de desperdicios de carne y víceras, mayormente de ganado vacuno. Este tipo de grasa se caracteriza por una mayor uniformidad, además de presentar un alto punto de fusión (>40°C) y un menor contenido de humedad e impurezas (<1,5%) así como de AGL, en comparación con otras fuentes de grasas (Brandt y Anderson, 1990; Zinn y Plascencia, 2004b).
Grasas mezcladas
Las grasas mezcladas son mezclas con diferentes proporciones de grasas de origen animal, aceites vegetales, así como aceites acidulados y subproductos de refinería. De la misma forma que la grasa amarilla, las mezclas no son uniformes en su composición; de hecho, su composición es aún más variable, por lo que es difícil caracterizarla de una manera generalizada. Aún así, comparada con la grasa amarilla, es de apariencia más oscura y con un contenido mayor de AGL y materia insaponificable, tendiendo a poseer un valor de yodo más alto. Las características típicas de calidad para esta fuente de grasa son 90% mínimo de ácidos grasos totales (AGT) y niveles máximos de 50% de AGL, 3,5% de insaponificables, 1,5% de humedad y 1% de impurezas (Zinn, 1989a).
Extractos de jabón y otras fuentes grasas altas en AGL
Los extractos de jabón son subproductos resultantes de los procesos de la refinación de aceites comestibles. La composición de ácidos grasos es muy similar a la fuente original, pero con más contenido de AGL (>50%). Otra fuente de grasa alta en AGL es la grasa denominada "grasa de trampa" (griddle grease), la cual es obtenida en las trampas del desagüe de cocinas de cafeterías y restaurantes. Este tipo de grasa se ha incrementado en el mercado en los últimos años como resultado de recientes regulaciones medioambientales que indican que la grasa que se vierte al caño por error debe ser recuperada y reciclada. La composición es muy similar a la grasa amarilla, pero contiene tres veces más AGL (Plascencia et al., 1999).
Factores que Inciden en la Digestibilidad de las Grasas en Bovinos de Engorda
El asignar individualmente un valor energético por fuente o tipo de grasa es difícil. Una revisión de estudios de digestibilidad y comportamiento muestra una variación sustancial en los valores estimados de su contenido de energía. Por otra parte, el principal problema cuando son comparadas distintas fuentes de grasas, es que las grasas adicionadas en las dietas para rumiantes generalmente no exceden del 6% de la materia seca y la precisión obtenida en esos estudios no permite detectar diferencias tan pequeñas (menos del 10%) en el valor nutricional de las grasas comparadas. De cualquier forma, al comparar distintas fuentes, básicamente se comparan características tales como la cantidad de AGL, el grado de saturación y la proporción de insaturados:saturados. En ese sentido, se han realizado varias pruebas para comparar el valor nutricional de las diferentes características de las grasas utilizadas en la alimentación para ganado en engorda.
Ácidos grasos libres (AGL)
Los ácidos grasos libres son ácidos grasos no esterificados con glicerol. En grasas y aceites la presencia de niveles altos de AGL puede indicar un almacenamiento o manejo inapropiado de la grasa (Valenzuela, 1995). La hidrólisis puede ocurrir en forma de lipólisis enzimática durante el almacenamiento o previo al procesado para su obtención, o presentarse como resultado de una hidrólisis autocatalítica denominada rancidez oxidativa (Barreras-Arellano, 1998).
TABLA I / COMPOSICIÓN QUÍMICA PROMEDIO DE LAS PRINCIPALES GRASAS COMERCIALES UTILIZADAS EN ENGORDA DE GANADO EN CONFINAMIENTO
1 Zinn, 1992; Krehbiel et al., 1995; Plascencia et al., 1999; Zinn et al., 2000; Ramírez and Zinn, 2000.
2 Palmquist, 1991; Clary et al., 1993; Krehbiel et al., 1995; Elliot et al., 1997; Beam, 2000; Ramirez and Zinn, 2000.
3 Zinn, 1989a; Palmquist, 1991; Wu et al., 1991.
4 AFOA, 1999; Bock et al., 1991; Zinn, 1992.
5 Coppock and Wilks, 1991; Klusmeyer and Clark, 1991; Palmquist, 1991; Wu et al., 1991, 1993; Zinn y Plascencia, 1992.
El efecto del nivel de AGL en la dieta sobre el comportamiento productivo ha sido estudiado en la mayoría de las especies, especialmente en pollos de engorda. El crecimiento en pollos generalmente no es afectado por consumos de dietas con grasa que contengan niveles altos de AGL (Vila y Esteve-García., 1996). Sin embargo, existen indicios en rumiantes, en los cuales los AGL pueden ser menos digestibles que los triglicéridos. Czerkawski (1973) observó diferencias en la digestión del aceite de semilla de lino dependiendo si se agregaba a la dieta como triglicérido (85% digerido) o en forma de ácidos grasos libres (64% digerido). En contraste, Zinn (1989a, b) no detectó diferencias significativas al comparar grasa amarilla, que es una fuente baja en AGL (10%), con una mezcla de grasa animal-vegetal (50% AGL) las cuales fueron adicionadas en dietas de finalización para bovinos a niveles de 4 y 8%. El comportamiento productivo, el valor de EN estimada y la digestibilidad intestinal de los ácidos grasos fueron similares en ambas fuentes de grasa y no resultaron influenciados por los niveles de adición.
Considerando la composición química de los triglicéridos, el glicerol es un diluyente del contenido neto de energía, al incrementarse el contenido de ácidos libres en una fuente de grasa se espera que el contenido energético se aumente de la misma manera. En ese sentido, Plascencia et al. (1999) evaluaron la influencia de distintos niveles de AGL contenidos en la grasa amarilla adicionada a las dietas para bovinos de engorda sobre el comportamiento productivo y digestión de nutrientes. Los niveles probados fueron 42,0; 28,5 y 15% de AGL en una dieta que contuvo 5% de grasa amarilla adicionada y 79% de grano en hojuela (50:50 de cebada y maíz). Aún cuando no existieron diferencias en la digestibilidad postruminal de los AG entre los tratamientos, el incremento de contenido de AGL aumentó en forma lineal la ganancia diaria de peso, el consumo y la conversión alimenticia. Los resultados obtenidos por Plascencia et al. (1999) han sido utilizados como indicios para soportar la hipótesis del efecto de dilución por glicerol utilizado en no rumiantes (Hamilton, 2002). Sin embargo, una de las características de los lípidos que ingresan en el intestino en rumiantes es que en su gran mayoría (>85%) son en forma no esterificada, lo que debilita dicha teoría, al menos para esta especie. Una explicación más aceptable cuando se observan respuestas positivas con grasas de mayor contenido de AGL es que éstos inhiben la tasa de biohidrogenación ruminal (Noble et al., 1974), lo que aumenta el flujo al duodeno de ácidos grasos de mayor digestibilidad (insaturados). Sin embargo, esta teoría no ha sido plenamente confirmada.
Proporción de insaturados:saturados
Recientemente ha surgido la controversia acerca del efecto potencial de la proporción de ácidos insaturados:saturados contenidos en las grasas sobre su valor nutricional para bovinos en engorda. Aunque existe limitada información para ganado de engorda, estudios in vitro (Henderson, 1973; Maczulak et al., 1981) han demostrado que los ácidos grasos insaturados juegan un papel más activo en la inhibición de las bacterias ruminales, particularmente las celulolíticas. De los ácidos grasos insaturados evaluados, el oleico (C18:1) fue el mayor inhibidor. Considerando que las bacterias celulolíticas participan menos en la función digestiva del ganado con dietas de finalización, se ha pensado que los efectos de la proporción de ácidos insaturados serían limitados en esas condiciones. No obstante lo anterior, se han llevado a cabo varias pruebas de comportamiento y de digestión en bovinos de engorda alimentados con dietas altas en energía con la finalidad de comparar grasas con alto grado de saturación frente a aquellas de menor grado de saturación. Por ejemplo, Brandt y Anderson (1990) al comparar sebo de res y grasa amarilla observaron respuestas similares y positivas para ambas fuentes de grasa en un primer experimento, mientras que en un segundo experimento, el comportamiento en consumo y ganancia de peso de los novillos que recibieron las dietas que contenían grasa amarilla fueron marcadamente menores que con la dieta que contuvo sebo. Estos resultados son desconcertantes, considerando que en ambos experimentos el nivel de adición fue bajo (3,5%), y las dietas y fuentes de grasa similares. En otros casos, el ganado que consumió dietas con sebo obtuvo menores ganancias y conversión alimenticia que aquellos que consumieron grasa amarilla (Lofgreen, 1965; Huffman et al., 1992). Aún así, la mayoría de los estudios no han detectado diferencias entre ambas fuentes de grasa, aún cuando se han añadido en niveles de 6% o más (Robert y McKirdy, 1964; Zinn, 1989a, b, 1992; Plascencia y Zinn, 2001).
En general, con el incremento de saturación de una fuente de grasa en particular (por ejemplo mediante hidrogenación) disminuyen los efectos negativos sobre la fermentación ruminal, pero también se reduce la digestibilidad intestinal de los ácidos grasos. En ese sentido, se han demostrado disminuciones en la digestibilidad de los ácidos grasos de sebo de res desde 74% para sebo de res nativo a 37% para sebo altamente hidrogenado (Macleod y Buchannan-Smith, 1972; Elliott et al., 1999), y reducciones del 23% en la digestibilidad de la grasa amarilla cuando esta se ofrece en forma hidrogenada (Jenkins y Jenny, 1989). Esto no debe ser generalizado entre diferentes fuentes de grasa con grado distinto de saturación. Por ejemplo, Bock et al. (1991) no detectaron diferencias en la digestibilidad intestinal de los ácidos grasos (promedio 75%) del extracto de jabón de aceite de soja con una proporción de insaturados:saturados de 3,7 comparado con el sebo de res con una proporción de insaturados:saturados de 1,65 adicionados a una dieta de finalización que contenía 78% de trigo quebrado. De igual forma, Plascencia et al. (2001) no observaron diferencias en la utilización intestinal de ácidos grasos cuando compararon la adición de 5% de grasa amarilla o sebo vacuno en dietas de finalización. Adicionalmente, Palmquist (1991) no obtuvo diferencias en la digestibilidad de los ácidos grasos cuando comparó 5 fuentes de grasa con diferentes proporciones de insaturados:saturados.
La atenuación de los efectos de la proporción de insaturados:saturados contenidos en las grasas que comúnmente se adicionan a las dietas para rumiantes se debe principalmente al elevado grado de biohidrogenación que sufren los AG insaturados en su estancia en rumen (Jenkins, 1993). La biohidrogenación ruminal es el mecanismo mediante el cual los microorganismos ruminales saturan los AG insaturados C18 hasta esteárico (Polan et al., 1964), aumentando de esta manera la cantidad de AG saturados que llega al intestino. La biohidrogenación de los ácidos grasos insaturados, es un mecanismo para reducir la toxicidad de los mismos para las bacterias, ya que por sus características poseen una acción detergente para la membrana celular microbiana (Garnsworthy, 2002). Para que la biohidrogenación se realice es necesario que tenga lugar previamente la hidrólisis de los lípidos (triglicéridos, galactolípidos y fosfolípidos) a nivel ruminal, la cual permite la disponibilidad del grupo carboxilo necesario para el proceso (Demeyers y Henderickx, 1967). Esa situación se ve disminuida cuando los AG son consumidos como AGL. (Jenkins, 1993). La hidrólisis ocurre por acción de lipasas, galactosidasas y fosfolipasas producidas por bacterias ruminales, principalmente Anaerovibrio lipolityca y Butirivrio sp. (Yokohama y Johnson, 1988). Algunos de los factores de mayor importancia que afectan la tasa de biohidrogenación son el pH ruminal (Van Nevel y Demeyer, 1996), la población microbiana (Latham et al., 1972), la naturaleza de los lípidos consumidos (Byers y Shelling, 1988) y la tasa de pasaje (recambio ruminal de ácidos grasos; Harfoot y Hazlewood, 1997), entre otros. Aún cuando la tasa de biohidrogenación de C18:1 aparentemente no está influenciada por el nivel de ácidos grasos consumidos (Duckett et al., 2002), estudios realizados in vitro por Beam et al. (2000) indicaron que por cada unidad porcentual incrementada de C18:2 en la dieta, la tasa de biohidrogenación de ese ácido graso disminuía en 0,12%/h. Considerando que la lipolisis ruminal de los triglicéridos es intensa (>90%/h; Inming et al., 1993), el grado de biohidrogenación de los ácidos grasos insaturados es también intensa y se encuentra en un rango entre 60 y 93%, con una media de 70% para grasas no protegidas (Hawke y Silcock, 1970; Wu et al., 1991, Pantoja et al., 1996, Plascencia et al., 1999; Zinn et al., 2000; Wachira et al., 2000) y de 47 a 57% de las grasas protegidas (Klusmeyer y Clark, 1991; Wu et al., 1991; Doreau et al., 1999; Zinn et al., 2000). Lo anterior se refleja en forma directa en la proporción y tipo de ácido graso que fluye al duodeno. Por ejemplo, Wu et al. (1991), alimentando a vacas con una mezcla animal-vegetal (59% de insaturados), notaron que el consumo de ácido linoleico aumentó de 171 a 296g/d; sin embargo, el flujo a duodeno aumentó sólo de 45 a 54g/d. De igual forma, en novillos alimentados con dietas adicionadas con extractos de jabón de aceite de soja (84% insaturados), el consumo de C18:1 aumentó de 38 a 99g/d, mientras que el flujo a duodeno sólo fue de 19 a 27g/d (Bock et al., 1991). Lo anterior demuestra que las diferencias entre las proporciones en los AG existentes entre las distintas fuentes de grasa consumidas disminuyen cuando éstos llegan al intestino, de tal forma que la proporción de insaturados:saturados en el alimento tiene una menor relevancia para rumiantes comparado con las especies no rumiantes (Cera et al., 1989; Vila y Esteve-García, 1996).
Basado en lo anterior, un punto abordado recientemente en rumiantes es la proporción de saturados (C16:0 y C18:0) y la cantidad de insaturados que ingresan al duodeno. La mezcla de grasas saturadas (sebo) con insaturadas (aceite de soja) ha resultado en efectos asociativos positivos en el valor nutricional de las grasas suplementarias para ganado de engorda (Brandt y Anderson, 1990). Esta respuesta es atribuida a que potencialmente los ácidos grasos insaturados pueden aumentar la absorción intestinal de los ácidos grasos saturados (Elliott et al., 1997; Zinn et al., 2000). Al respecto, Firkins y Eastridge (1994) demostraron que a una mayor proporción de C16:0 con respecto a C18:0 del total de AG saturados generalmente mejora la digestibilidad de los ácidos grasos, especialmente si el grado de insaturados de la grasa se incrementa. Zinn et al. (2000) observaron que al disminuir el índice de biohidrogenación mediante la protección de la grasa con una matriz de formaldehído-proteína mejoró la digestión intestinal de los AG totales (87,8 vs. 80,3%). Adicionalmente detectaron que por cada 1% de incremento en la proporción de C18:1 en relación al total de AG que llegan al intestino, la digestibilidad de C18:0 aumentaba en 1% (Digestibilidad de C18:0= 85,72+1,01P18:1-15,84FI; donde P18:1 es el C18:1 ingresando al intestino expresado como porcentaje del total de los AG que llegan al duodeno y FI es la cantidad diaria de AG consumidos expresados como g de AG/kg de PV, r2= 0,99). Apoyados en el concepto de que la digestión intestinal de los ácidos grasos saturados disminuye a medida que la longitud de su cadena aumenta (Steele y Moore, 1968), es razonable esperar que el ácido palmítico (C16:0) tiene condiciones más favorables para mostrar un efecto sinérgico mayor con los ácidos grasos insaturados a nivel postruminal que el ácido esteárico (C18:0). En ese sentido, Plascencia et al. (2003a) evaluaron una fuente de grasa rica en palmítico (>97%) protegida con formaldehído mezclada al 50% con grasa amarilla, la cual es rica en insaturados (>65%), añadida a un nivel de 5% de la dieta. La combinación resultó en una mejora de la digestibilidad intestinal de los ácidos grasos de 5% cuando se comparó con las fuentes de grasas originales.
Métodos de adición
Debido a sus características físicas las grasas tienden a formar una capa o cubierta en las partículas alimenticias, principalmente fibra (Devendra y Lewis, 1974). De hecho aproximadamente un 80% del total de lípidos en rumen está en forma asociada a partículas (McAllan et al., 1983) y de 40 a 75% de las bacterias está adherido a las partículas alimenticias (Owens y Goetsch, 1988). Esto hace que por sus propiedades hidrófobas las grasas por cubrimiento físico puedan ejercer un efecto inhibitorio a la acción enzimática bacteriana interfiriendo en los procesos normales de fermentación (Devendra y Lewis, 1974) y afectando, por efectos asociativos, el valor nutricional de las grasas alimenticias. En ese sentido, se han conducido una serie de estudios con la finalidad de evaluar si el método de adición de grasa puede influir en su valor nutricional. Zinn et al. (1998) evaluaron la adición de grasa amarilla, bien sea añadida primeramente al grano (maíz en hojuela) y el producto resultante mezclado con el resto de los ingredientes de la dieta, o bien la grasa amarilla era añadida al final del mezclado. En ambas situaciones la cantidad final de grasa en la dieta fue de 5%. No se detectaron diferencias entre los métodos de adición sobre los parámetros digestivos o el comportamiento productivo del ganado.
En virtud de que el punto de fusión de las grasas afecta su grado de lipólisis (Beam et al., 2000), una fuente de grasa de mayor punto de fusión podría tener un efecto diferente en cuanto a su método de adición. Para tal efecto Plascencia et al. (2001), mediante una prueba de digestión y metabolismo evaluaron, empleando el mismo esquema de tratamientos utilizados por el estudio de Zinn et al. (1998), si el método de adición puede ser afectado por el punto de fusión de la fuente de grasa utilizada (sebo vs. grasa amarilla). Al igual que el estudio de Zinn et al. (1998), no existieron efectos de los tratamientos sobre el sitio o grado de digestión de los diferentes componentes de la dieta. La digestión intestinal de la grasa promedió 69,3%. El realizar mezcla de grasa-forraje también ha sido evaluado en dietas de finalización para bovinos en engorda. La mezcla de 80% de alfalfa con 20% de grasa fue utilizada en 15, 30 y 45% en dietas de finalización. La mezcla al nivel de 15% contuvo 71% de trigo en hojuela y en los niveles de 30 y 45% fue incorporada sustituyendo directamente al trigo en hojuela. El sustituir el grano con la mezcla alfalfa-grasa no afectó el consumo o la ganancia diaria; sin embargo, la EN de la dieta disminuyó en forma lineal al aumentar la proporción de la mezcla en la dieta. De igual manera, la digestibilidad intestinal de los ácidos grasos disminuyó a medida que se aumentaba la mezcla en la ración. Se concluyó que la cantidad de grasa consumida fue el factor principal que limitó el contenido de EN y la digestibilidad de los AG, mientras que el método de adición no tuvo un papel relevante en los resultados (Plascencia y Zinn, 2002).
Con la finalidad de comprobar los resultados obtenidos en los estudios anteriores se realizó un experimento de comportamiento productivo con 216 novillos (Zinn y Plascencia, 2004a). Se evaluaron tres métodos de adición de grasa (ácidos grasos de sebo añadido bien en el grano, en el forraje o en la ración completa) con tres niveles de inclusión (3, 6 y 9%). El aumento del nivel de grasa en la dieta disminuyó el consumo, la ganancia diaria de peso y la EN de la grasa en forma lineal. Nuevamente el nivel de consumo fue el factor primordial, mientras que el método de adición no mostró efectos asociativos sobre el valor nutricional de la grasa adicionada.
Nivel de inclusión
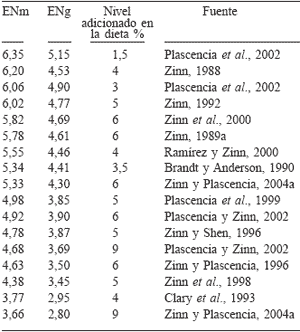
Las recomendaciones para el uso de grasas alimenticias para dietas de rumiantes indican que éstas no deben exceder el 5% de la dieta, puesto que se han observado efectos detrimentales sobre el consumo y la eficiencia alimenticia cuando la grasa se incluye en niveles superiores (Haaland et al., 1981; Ngidi et al., 1990; Zinn, 1994). Sin embargo, las restricciones prácticas para su óptima utilización no han sido aún resueltas, ya que se han registrado casos negativos en comportamiento productivo con niveles de inclusión igual o menor al 3% (Hatch et al., 1972; Krehbiel et al., 1995), mientras que niveles de 8% han conducido a ganancias y conversiones superiores con relación a animales no suplementados (Zinn, 1989a). Lo anterior se refleja en una variabilidad del valor nutricional observado para la energía neta (EN) de la grasa que oscila desde 3,77 y 2,95 (Clary et al., 1993) hasta 6,35 y 5,15Mcal·kg-1 (Plascencia et al., 2002) de EN de mantenimiento y EN de ganancia, respectivamente, tal como se aprecia en la Tabla II.
Aún así, una respuesta generalizada cuando se aumenta el nivel de grasa en la dieta es la disminución del valor energético de la grasa. Por ejemplo, Zinn y Plascencia (2004a) informan que al aumentar el nivel de grasa de 3 a 9% en una dieta de finalización para novillos, se observaron disminuciones en el consumo, ganancia diaria y conversión alimenticia. La EN de la dieta resultó ser 103% para el nivel de inclusión de 3% y descendió a 90% cuando la grasa fue añadida en un 9% en la dieta. La EN calculada de la grasa declinó de 6,4 a 3,44Mcal·kg-1. La razón para lo anterior ha sido atribuido principalmente a variaciones en la digestión intestinal de lípidos (Wu et al., 1991; Zinn, 1994). El valor de la energía bruta (EB) para las grasas es de 9,4Kcal·g-1; sin embargo, el valor combustible "utilizable" por los tejidos debe considerarse a partir de su digestibilidad. Las grasas de grado alimenticio contienen aproximadamente 90% de ácidos grasos totales (AFOA, 1999), y éstos representan casi el 100% de su contenido energético (Zinn, 1989a). Por lo tanto, el valor energético de las grasas está supeditado a la digestibilidad de sus ácidos grasos la cual en bovinos es aproximadamente del 77% (Tabla III). Bauchart (1993) indica un 80% de digestibilidad para los AG saturados y de 93% para los insaturados cuando se consumen dietas con moderado contenido de lípidos (2-3%). La elevada capacidad de los rumiantes para digerir AG saturados, con respecto a los no rumiantes, se atribuye a las condiciones de pH duodenal (2-2,5, Christiansen y Webb, 1990), la proporción de fosfatidilcolina (80%) con alto contenido de oleico (Christie, 1973), así como por la elevada proporción de taurocolatos (Peric-Golia y Socic, 1968) de las sales biliares. Por otra parte, el concepto generalizado de que la longitud de la cadena en los AG saturados afecta negativamente a su digestibilidad (Steele y Moore, 1968) no parece ser de importancia en rumiantes cuando los consumos de lípidos son moderados (Weisjberg et al., 1992). Sin embargo, a altos consumos, la digestibilidad de C18:0 es 5 a 8% menor en relación a C16:0 (Bauchart, 1993). Como se mencionó anteriormente, la digestibilidad de los ácidos grasos insaturados es mayor que la de los saturados y no parecen ser afectados aún con altos consumos de grasa (Ávila et al., 2000; Enjalbert et al., 2000; Plascencia et al., 2003b). Existen informes que indican que el ácido linolénico es absorbido menos eficientemente que el palmítico (Doreau y Ferlay, 1994); sin embargo, esos resultados son aparentes, ya que esas diferencias se atribuyen a problemas metodológicos en la determinación de C18:3, dada su baja concentración (<2%) en el flujo duodenal.
Lo anterior demuestra, en gran medida, que el principal factor que afecta la digestibilidad de la grasa alimenticia en rumiantes es su nivel de consumo (Palmquist y Conrad, 1980; Ngidi et al., 1990; Coppock y Wilks, 1991; Palmquist, 1991; Khorasani et al., 1992; Zinn, 1992, 1994; Pylot et al., 2000). Por ejemplo, Zinn (1989b) observó una disminución en forma lineal (83, 81 y 74%) de la digestibilidad intestinal de los ácidos grasos cuando se aumentó el nivel de inclusión de grasa amarilla en la dieta de 0 a 8%, calculando una disminución de la digestibilidad de la grasa en 3,4% por cada unidad porcentual por encima de 4% de inclusión. Similares a lo anterior son los resultados de Palmquist (1991), quien observó que la digestibilidad de los ácidos grasos disminuía en 2,2% por cada 100g de ácidos grasos consumidos. Adicionalmente, Palmquist y Conrad (1980) indican que a niveles moderados de inclusión (por debajo de 5%) la digestibilidad verdadera de la grasa es aproximadamente de 80%, mas si se agrega la grasa por encima de ese nivel se digiere menos eficientemente (56%). Por otra parte, Wu et al. (1991) determinaron que la reducción de la digestibilidad total de ácidos grasos se debía principalmente al efecto negativo de la digestibilidad del ácido esteárico (C18:0), ya que registraron una disminución en su digestibilidad de un 21% cuando la grasa suplementada fue incrementada en la dieta del 3 al 6% (72 y 59%, respectivamente). Estos datos son muy similares al 75% y 59% de digestibilidad para C18:0 obtenidos por Zinn (1992) en dietas con 90% de concentrados suplementados con 0% y 6% de grasa animal.
TABLA II / COMPARACIÓN DEL CONTENIDO DE ENERGIA NETA* DE MANTENIMIENTO Y DE GANANCIA DE GRASAS ALIMENTICIAS ESTIMADO POR MEDIO DE LA TÉCNICA DE REEMPLAZO EN NOVILLOS CON DIETAS DE FINALIZACIÓN
* Mcal·kg-1.
TABLA III / DIGESTIBILIDAD INTESTINAL PARA RUMIANTES DE LAS DISTINTAS FUENTES DE GRASA
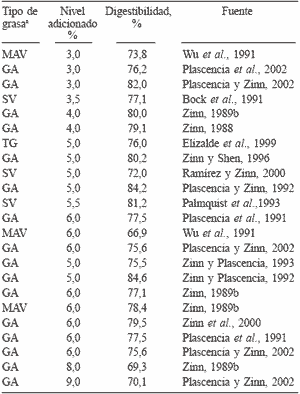
a MAV: mezcla animal-vegetal, GA: grasa amarilla, TG: triglicéridos, SV: sebo vacuno.
Los estudios anteriores muestran concordancia en que a mayor nivel de inclusión menor es la eficiencia de utilización intestinal de los ácidos grasos, principalmente los saturados (C16:0 y C18:0). Sin embargo, no existe concordancia respecto a qué porcentaje de inclusión permite el máximo de utilización de grasa sin afectar negativamente su digestibilidad, y por lo tanto su contenido de EN. Aún así, las recomendaciones prácticas actuales indican que la inclusión no exceda del 5% de la dieta. Aunque Doreau y Ferlay (1994), al relacionar la digestibilidad intestinal con el consumo de grasa de 13 experimentos, no encontraron relación entre el nivel de grasa en la dieta y la digestión de los ácidos grasos, es importante hacer notar que la relación establecida por esos investigadores fue la de la concentración de lípidos (g·kg-1 de MS) frente a digestibilidad expresada en porcentaje. Esta relación puede confundir los efectos, ya que indica sólo la concentración de lípidos en la dieta y no la cantidad total de lípidos consumidos. Respecto a lo anterior, Brandt et al. (1992) indican que mucha de la inconsistencia en la respuesta del comportamiento productivo del ganado cuando consume dietas suplementadas con grasa puede estar más asociada con la cantidad total de lípidos consumidos que con el porcentaje de inclusión.
Al evaluar el efecto del nivel de consumo de ácidos grasos sobre el valor nutricional de las grasas relacionándolo con su digestión intestinal, Plascencia et al. (2003b) observaron una correlación negativa de la cantidad de AG consumidos (g·kg-1 de peso corporal) frente a su digestibilidad intestinal. Combinando los resultados obtenidos en ese estudio con otros 7 estudios previos se generó la ecuación de regresión (Figura 1)
y= 87,56 - 8,591x (R2= 0,89, n=25)
donde y: digestibilidad intestinal, y X: ácidos grasos consumidos por día (g·kg-1 de peso corporal). Esta regresión implica que para no afectar a la digestibilidad intestinal de los AG, el consumo diario de ácidos grasos totales debe ser menor a 0,96g de AGT por kg de peso corporal.
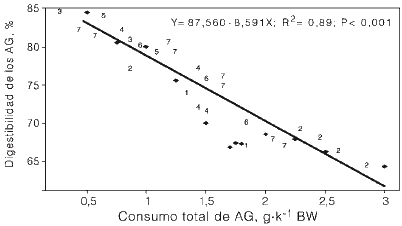
Figura 1. Influencia del nivel de consumo de ácidos grasos (AG)sobre su digestión intestinal.
Lo anterior indica que la principal limitante en la digestibilidad sucede cuando la cantidad de los lípidos que llegan al duodeno sobrepasa la capacidad enzimática intestinal y, por consiguiente, su absorción (Bauchart, 1993). Los ácidos grasos que llegan al intestino en rumiantes son altamente saturados (>65%) y están asociados a partículas (Wu et al., 1991; Pantoja et al., 1996; Plascencia et al., 1999). Los AG saturados son físicamente menos voluminosos y más viscosos que los insaturados, lo cual dificulta la formación de la fase micelar (Moore y Christie, 1984) y la tasa de absorción, puesto que se ha comprobado que la absorción de los AG saturados es más lenta (Doreau y Ferlay, 1994). Debe considerarse que la forma de evaluar la digestibilidad se basa no sólo en el potencial de digestión del nutriente en el organismo, sino también en su potencial de absorción en el tracto gastrointestinal, y es importante que se considere esta limitante cuando se presentan altos consumos de lípidos. La disminución de la digestibilidad de los AG saturados puede explicar de 85 a 100% la variación del valor nutricional observado para las grasas adicionadas a las dietas para rumiantes (Pantoja et al., 1995; Ramírez y Zinn, 2000; Ávila et al., 2000; Enjalbert et al., 2000) y esto puede estar relacionado con una posible emulsificación incompleta de los AG saturados en el intestino delgado como resultado de una producción inadecuada de jugo biliar con relación a la cantidad presente de estos ácidos grasos que se observa como resultado del consumo de dietas con alto contenido de grasa (Palmquist, 1991; Xu et al., 1998). En ese sentido, en aves se ha demostrado deficiencias en la utilización adecuada de las grasas alimenticias por insuficiencia de jugo biliar. Atteh y Lesson (1985) observaron un incremento del 16% en la digestión intestinal de una mezcla de palmítico:oleico cuando se añadió 0,2% de ácido cólico a la dieta, lo que se tradujo en una mejora de un 14% en la energía digestible de la dieta. Considerando que en rumiantes la producción de jugo biliar oscila de 0,5 a 1,45ml·kg-1 de peso (Harrison, 1962; Moore y Christie, 1984; Merchen, 1988), para un bovino de 200kg de peso vivo la producción biliar varía desde 2400 hasta 6960ml/día. Sin embargo, las diferencias detectadas en la producción biliar evaluada por métodos de cateterización del conducto biliar utilizando técnicas de recolecciones repetidas de muestras, pueden ser debidas a la disminución de la resorción de jugos biliares en las partes distales del intestino (reflejo enterocólico; Hurwitz et al., 1973) que afectan la producción biliar a corto plazo, a la disminución del flujo biliar debido a irritación directa en el conducto biliar (Harrison, 1995), a la inexactitud en la técnica de recolección de la totalidad jugo biliar (Merchen, 1988), o bien a características propias de la dieta. Con respecto a este último punto, aún cuando se ha determinado que el flujo biliar en rumiantes tiende a ser independiente de la hora del día o la frecuencia alimenticia (Symonds et al., 1982), es importante considerar a la cantidad de ácidos grasos y a la proporción de insaturados:saturados que ingresan al duodeno como una variable potencial sobre la producción biliar, ya que éstos tienen un efecto directo sobre la producción de gastrina y colecistoquinina, ambos elementos involucrados en el mecanismo de flujo biliar al duodeno (Studzinsky y Bobowiec, 1979; Kato et al., 1991).
TABLA IV / PRODUCCIÓN BILIAR Y DIGESTIÓN POSRUMINAL DE LOS ÁCIDOS GRASOSTOTALES EN NOVILLOS* CONSUMIENDO DIETAS CON 3 NIVELES DEGRASA SUPLEMENTARIA
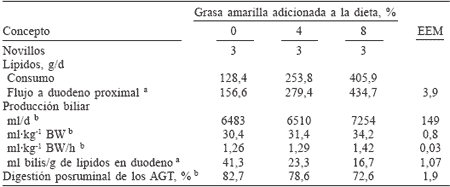
* De 215kg de peso.
a Efecto lineal del nivel de grasa, P<0,01.
b Efecto lineal del nivel de grasa, P<0,10.
Se ha argumentado que el grado de formación de micelas, primer paso para la absorción de lípidos, puede variar con el tipo de dieta, lo que podría ocurrir sólo si se aumenta la producción biliar o la proporción de sales biliares:lípidos, o bien si la proporción de ácidos grasos insaturados:saturados en el duodeno aumenta. Sin embargo, existen estudios en los que se ha probado el uso de emulsificantes, principalmente lecitina, combinados con la grasa alimenticia en los cuales no se han obtenido beneficios en la digestión postruminal de los ácidos grasos (Zinn, 1989b; Jenkins y Fotohui, 1990; Plascencia et al., 1991). Lo anterior puede explicarse por la alta hidrólisis ruminal de la lecitina, lo que evita que llegue con sus características emulsificantes al intestino (Jenkins, 1990). Un indicio de la limitada capacidad de emulsificación a nivel intestinal en rumiantes lo muestra el estudio de Børsting et al. (1992), quienes indican coeficientes de digestión de 92% para C18:0 cuando las vacas consumieron dietas con grasas vegetales emulsificadas y protegidas contra la hidrólisis ruminal mediante una matriz de formaldehído-caseína. Esos investigadores concluyeron que los bajos coeficientes de digestión observados en la mayoría de los estudios para C18:0 con altos consumos de lípidos son debidos principalmente a una pobre emulsificación más que a un inefectivo sistema de lipasas. En ese sentido, Plascencia et al. (2004) relacionaron la producción biliar con la digestión postruminal de los AG en novillos que consumieron dietas con tres distintos niveles de grasa amarilla. La producción biliar promedió 32ml·kg-1 de PV y se incrementó (efecto lineal) a medida que se aumentó el flujo de lípidos al duodeno. Sin embargo, considerada como proporción del flujo de lípidos (ml de bilis por g de lípido que ingresa al duodeno proximal) la producción biliar disminuyó linealmente de 23,4 a 16,7ml de bilis/g de lipidos en duodeno (Tabla IV). La mayor digestibilidad para AG se observó en una relación de 40ml de bilis/ g de lípidos en duodeno y la producción biliar explicó en 69% la variación en la utilización de AG en ese experimento (Figura 2).
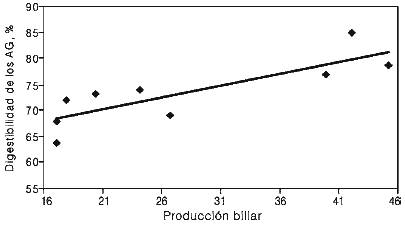
Figura 2. Influencia de la producción biliar (ml de bilis por g delípidos en duodeno) sobre la digestión intestinal de los AG.
Conclusiones
En relación a las grasas libres, son diversas las fuentes de grasa que son utilizadas en la alimentación de bovinos de engorda. Éstas difieren principalmente en contenido de impurezas, de ácidos grasos libres (AGL) y en el grado de saturación (Tabla I). A continuación se describen brevemente las fuentes de grasas más comúnmente utilizadas.
Grasa amarilla
El término de "amarilla" se debe a su apariencia. También se le conoce como grasa de restaurante o grasa de cocina, ya que su origen es de cualquier combinación de los desperdicios o sobrantes de grasas y aceites colectados en cafeterías, restaurantes de comida rápida y panaderías. Como resultado de cocinar cada vez más con aceites vegetales, la mayor parte de grasa amarilla recobrada es de origen vegetal que ha sido parcialmente hidrogenizada para un mejor desempeño en el proceso de cocinado, de tal forma que la proporción de insaturados:saturados es de 2,6 aproximadamente (Zinn, 1988; Plascencia et al., 1991). Debido a la diversidad de sus fuentes, la grasa amarilla no es muy uniforme en su composición y puede variar de un área a otra, o de una planta a otra. De acuerdo a los parámetros establecidos por la Asociación Americana de Grasas y Aceites (AFOA, 1999), su punto de fusión debe ser menor a 40ºC y no debe contener más de 15% de AGL y un máximo de 2% de impurezas.
Sebo
El sebo o grasa animal es un subproducto derivado principalmente de desperdicios de carne y víceras, mayormente de ganado vacuno. Este tipo de grasa se caracteriza por una mayor uniformidad, además de presentar un alto punto de fusión (>40°C) y un menor contenido de humedad e impurezas (<1,5%) así como de AGL, en comparación con otras fuentes de grasas (Brandt y Anderson, 1990; Zinn y Plascencia, 2004b).
Grasas mezcladas
Las grasas mezcladas son mezclas con diferentes proporciones de grasas de origen animal, aceites vegetales, así como aceites acidulados y subproductos de refinería. De la misma forma que la grasa amarilla, las mezclas no son uniformes en su composición; de hecho, su composición es aún más variable, por lo que es difícil caracterizarla de una manera generalizada. Aún así, comparada con la grasa amarilla, es de apariencia más oscura y con un contenido mayor de AGL y materia insaponificable, tendiendo a poseer un valor de yodo más alto. Las características típicas de calidad para esta fuente de grasa son 90% mínimo de ácidos grasos totales (AGT) y niveles máximos de 50% de AGL, 3,5% de insaponificables, 1,5% de humedad y 1% de impurezas (Zinn, 1989a).
Extractos de jabón y otras fuentes grasas altas en AGL
Los extractos de jabón son subproductos resultantes de los procesos de la refinación de aceites comestibles. La composición de ácidos grasos es muy similar a la fuente original, pero con más contenido de AGL (>50%). Otra fuente de grasa alta en AGL es la grasa denominada "grasa de trampa" (griddle grease), la cual es obtenida en las trampas del desagüe de cocinas de cafeterías y restaurantes. Este tipo de grasa se ha incrementado en el mercado en los últimos años como resultado de recientes regulaciones medioambientales que indican que la grasa que se vierte al caño por error debe ser recuperada y reciclada. La composición es muy similar a la grasa amarilla, pero contiene tres veces más AGL (Plascencia et al., 1999).
Factores que Inciden en la Digestibilidad de las Grasas en Bovinos de Engorda
El asignar individualmente un valor energético por fuente o tipo de grasa es difícil. Una revisión de estudios de digestibilidad y comportamiento muestra una variación sustancial en los valores estimados de su contenido de energía. Por otra parte, el principal problema cuando son comparadas distintas fuentes de grasas, es que las grasas adicionadas en las dietas para rumiantes generalmente no exceden del 6% de la materia seca y la precisión obtenida en esos estudios no permite detectar diferencias tan pequeñas (menos del 10%) en el valor nutricional de las grasas comparadas. De cualquier forma, al comparar distintas fuentes, básicamente se comparan características tales como la cantidad de AGL, el grado de saturación y la proporción de insaturados:saturados. En ese sentido, se han realizado varias pruebas para comparar el valor nutricional de las diferentes características de las grasas utilizadas en la alimentación para ganado en engorda.
Ácidos grasos libres (AGL)
Los ácidos grasos libres son ácidos grasos no esterificados con glicerol. En grasas y aceites la presencia de niveles altos de AGL puede indicar un almacenamiento o manejo inapropiado de la grasa (Valenzuela, 1995). La hidrólisis puede ocurrir en forma de lipólisis enzimática durante el almacenamiento o previo al procesado para su obtención, o presentarse como resultado de una hidrólisis autocatalítica denominada rancidez oxidativa (Barreras-Arellano, 1998).
TABLA I / COMPOSICIÓN QUÍMICA PROMEDIO DE LAS PRINCIPALES GRASAS COMERCIALES UTILIZADAS EN ENGORDA DE GANADO EN CONFINAMIENTO
1 Zinn, 1992; Krehbiel et al., 1995; Plascencia et al., 1999; Zinn et al., 2000; Ramírez and Zinn, 2000.
2 Palmquist, 1991; Clary et al., 1993; Krehbiel et al., 1995; Elliot et al., 1997; Beam, 2000; Ramirez and Zinn, 2000.
3 Zinn, 1989a; Palmquist, 1991; Wu et al., 1991.
4 AFOA, 1999; Bock et al., 1991; Zinn, 1992.
5 Coppock and Wilks, 1991; Klusmeyer and Clark, 1991; Palmquist, 1991; Wu et al., 1991, 1993; Zinn y Plascencia, 1992.
El efecto del nivel de AGL en la dieta sobre el comportamiento productivo ha sido estudiado en la mayoría de las especies, especialmente en pollos de engorda. El crecimiento en pollos generalmente no es afectado por consumos de dietas con grasa que contengan niveles altos de AGL (Vila y Esteve-García., 1996). Sin embargo, existen indicios en rumiantes, en los cuales los AGL pueden ser menos digestibles que los triglicéridos. Czerkawski (1973) observó diferencias en la digestión del aceite de semilla de lino dependiendo si se agregaba a la dieta como triglicérido (85% digerido) o en forma de ácidos grasos libres (64% digerido). En contraste, Zinn (1989a, b) no detectó diferencias significativas al comparar grasa amarilla, que es una fuente baja en AGL (10%), con una mezcla de grasa animal-vegetal (50% AGL) las cuales fueron adicionadas en dietas de finalización para bovinos a niveles de 4 y 8%. El comportamiento productivo, el valor de EN estimada y la digestibilidad intestinal de los ácidos grasos fueron similares en ambas fuentes de grasa y no resultaron influenciados por los niveles de adición.
Considerando la composición química de los triglicéridos, el glicerol es un diluyente del contenido neto de energía, al incrementarse el contenido de ácidos libres en una fuente de grasa se espera que el contenido energético se aumente de la misma manera. En ese sentido, Plascencia et al. (1999) evaluaron la influencia de distintos niveles de AGL contenidos en la grasa amarilla adicionada a las dietas para bovinos de engorda sobre el comportamiento productivo y digestión de nutrientes. Los niveles probados fueron 42,0; 28,5 y 15% de AGL en una dieta que contuvo 5% de grasa amarilla adicionada y 79% de grano en hojuela (50:50 de cebada y maíz). Aún cuando no existieron diferencias en la digestibilidad postruminal de los AG entre los tratamientos, el incremento de contenido de AGL aumentó en forma lineal la ganancia diaria de peso, el consumo y la conversión alimenticia. Los resultados obtenidos por Plascencia et al. (1999) han sido utilizados como indicios para soportar la hipótesis del efecto de dilución por glicerol utilizado en no rumiantes (Hamilton, 2002). Sin embargo, una de las características de los lípidos que ingresan en el intestino en rumiantes es que en su gran mayoría (>85%) son en forma no esterificada, lo que debilita dicha teoría, al menos para esta especie. Una explicación más aceptable cuando se observan respuestas positivas con grasas de mayor contenido de AGL es que éstos inhiben la tasa de biohidrogenación ruminal (Noble et al., 1974), lo que aumenta el flujo al duodeno de ácidos grasos de mayor digestibilidad (insaturados). Sin embargo, esta teoría no ha sido plenamente confirmada.
Proporción de insaturados:saturados
Recientemente ha surgido la controversia acerca del efecto potencial de la proporción de ácidos insaturados:saturados contenidos en las grasas sobre su valor nutricional para bovinos en engorda. Aunque existe limitada información para ganado de engorda, estudios in vitro (Henderson, 1973; Maczulak et al., 1981) han demostrado que los ácidos grasos insaturados juegan un papel más activo en la inhibición de las bacterias ruminales, particularmente las celulolíticas. De los ácidos grasos insaturados evaluados, el oleico (C18:1) fue el mayor inhibidor. Considerando que las bacterias celulolíticas participan menos en la función digestiva del ganado con dietas de finalización, se ha pensado que los efectos de la proporción de ácidos insaturados serían limitados en esas condiciones. No obstante lo anterior, se han llevado a cabo varias pruebas de comportamiento y de digestión en bovinos de engorda alimentados con dietas altas en energía con la finalidad de comparar grasas con alto grado de saturación frente a aquellas de menor grado de saturación. Por ejemplo, Brandt y Anderson (1990) al comparar sebo de res y grasa amarilla observaron respuestas similares y positivas para ambas fuentes de grasa en un primer experimento, mientras que en un segundo experimento, el comportamiento en consumo y ganancia de peso de los novillos que recibieron las dietas que contenían grasa amarilla fueron marcadamente menores que con la dieta que contuvo sebo. Estos resultados son desconcertantes, considerando que en ambos experimentos el nivel de adición fue bajo (3,5%), y las dietas y fuentes de grasa similares. En otros casos, el ganado que consumió dietas con sebo obtuvo menores ganancias y conversión alimenticia que aquellos que consumieron grasa amarilla (Lofgreen, 1965; Huffman et al., 1992). Aún así, la mayoría de los estudios no han detectado diferencias entre ambas fuentes de grasa, aún cuando se han añadido en niveles de 6% o más (Robert y McKirdy, 1964; Zinn, 1989a, b, 1992; Plascencia y Zinn, 2001).
En general, con el incremento de saturación de una fuente de grasa en particular (por ejemplo mediante hidrogenación) disminuyen los efectos negativos sobre la fermentación ruminal, pero también se reduce la digestibilidad intestinal de los ácidos grasos. En ese sentido, se han demostrado disminuciones en la digestibilidad de los ácidos grasos de sebo de res desde 74% para sebo de res nativo a 37% para sebo altamente hidrogenado (Macleod y Buchannan-Smith, 1972; Elliott et al., 1999), y reducciones del 23% en la digestibilidad de la grasa amarilla cuando esta se ofrece en forma hidrogenada (Jenkins y Jenny, 1989). Esto no debe ser generalizado entre diferentes fuentes de grasa con grado distinto de saturación. Por ejemplo, Bock et al. (1991) no detectaron diferencias en la digestibilidad intestinal de los ácidos grasos (promedio 75%) del extracto de jabón de aceite de soja con una proporción de insaturados:saturados de 3,7 comparado con el sebo de res con una proporción de insaturados:saturados de 1,65 adicionados a una dieta de finalización que contenía 78% de trigo quebrado. De igual forma, Plascencia et al. (2001) no observaron diferencias en la utilización intestinal de ácidos grasos cuando compararon la adición de 5% de grasa amarilla o sebo vacuno en dietas de finalización. Adicionalmente, Palmquist (1991) no obtuvo diferencias en la digestibilidad de los ácidos grasos cuando comparó 5 fuentes de grasa con diferentes proporciones de insaturados:saturados.
La atenuación de los efectos de la proporción de insaturados:saturados contenidos en las grasas que comúnmente se adicionan a las dietas para rumiantes se debe principalmente al elevado grado de biohidrogenación que sufren los AG insaturados en su estancia en rumen (Jenkins, 1993). La biohidrogenación ruminal es el mecanismo mediante el cual los microorganismos ruminales saturan los AG insaturados C18 hasta esteárico (Polan et al., 1964), aumentando de esta manera la cantidad de AG saturados que llega al intestino. La biohidrogenación de los ácidos grasos insaturados, es un mecanismo para reducir la toxicidad de los mismos para las bacterias, ya que por sus características poseen una acción detergente para la membrana celular microbiana (Garnsworthy, 2002). Para que la biohidrogenación se realice es necesario que tenga lugar previamente la hidrólisis de los lípidos (triglicéridos, galactolípidos y fosfolípidos) a nivel ruminal, la cual permite la disponibilidad del grupo carboxilo necesario para el proceso (Demeyers y Henderickx, 1967). Esa situación se ve disminuida cuando los AG son consumidos como AGL. (Jenkins, 1993). La hidrólisis ocurre por acción de lipasas, galactosidasas y fosfolipasas producidas por bacterias ruminales, principalmente Anaerovibrio lipolityca y Butirivrio sp. (Yokohama y Johnson, 1988). Algunos de los factores de mayor importancia que afectan la tasa de biohidrogenación son el pH ruminal (Van Nevel y Demeyer, 1996), la población microbiana (Latham et al., 1972), la naturaleza de los lípidos consumidos (Byers y Shelling, 1988) y la tasa de pasaje (recambio ruminal de ácidos grasos; Harfoot y Hazlewood, 1997), entre otros. Aún cuando la tasa de biohidrogenación de C18:1 aparentemente no está influenciada por el nivel de ácidos grasos consumidos (Duckett et al., 2002), estudios realizados in vitro por Beam et al. (2000) indicaron que por cada unidad porcentual incrementada de C18:2 en la dieta, la tasa de biohidrogenación de ese ácido graso disminuía en 0,12%/h. Considerando que la lipolisis ruminal de los triglicéridos es intensa (>90%/h; Inming et al., 1993), el grado de biohidrogenación de los ácidos grasos insaturados es también intensa y se encuentra en un rango entre 60 y 93%, con una media de 70% para grasas no protegidas (Hawke y Silcock, 1970; Wu et al., 1991, Pantoja et al., 1996, Plascencia et al., 1999; Zinn et al., 2000; Wachira et al., 2000) y de 47 a 57% de las grasas protegidas (Klusmeyer y Clark, 1991; Wu et al., 1991; Doreau et al., 1999; Zinn et al., 2000). Lo anterior se refleja en forma directa en la proporción y tipo de ácido graso que fluye al duodeno. Por ejemplo, Wu et al. (1991), alimentando a vacas con una mezcla animal-vegetal (59% de insaturados), notaron que el consumo de ácido linoleico aumentó de 171 a 296g/d; sin embargo, el flujo a duodeno aumentó sólo de 45 a 54g/d. De igual forma, en novillos alimentados con dietas adicionadas con extractos de jabón de aceite de soja (84% insaturados), el consumo de C18:1 aumentó de 38 a 99g/d, mientras que el flujo a duodeno sólo fue de 19 a 27g/d (Bock et al., 1991). Lo anterior demuestra que las diferencias entre las proporciones en los AG existentes entre las distintas fuentes de grasa consumidas disminuyen cuando éstos llegan al intestino, de tal forma que la proporción de insaturados:saturados en el alimento tiene una menor relevancia para rumiantes comparado con las especies no rumiantes (Cera et al., 1989; Vila y Esteve-García, 1996).
Basado en lo anterior, un punto abordado recientemente en rumiantes es la proporción de saturados (C16:0 y C18:0) y la cantidad de insaturados que ingresan al duodeno. La mezcla de grasas saturadas (sebo) con insaturadas (aceite de soja) ha resultado en efectos asociativos positivos en el valor nutricional de las grasas suplementarias para ganado de engorda (Brandt y Anderson, 1990). Esta respuesta es atribuida a que potencialmente los ácidos grasos insaturados pueden aumentar la absorción intestinal de los ácidos grasos saturados (Elliott et al., 1997; Zinn et al., 2000). Al respecto, Firkins y Eastridge (1994) demostraron que a una mayor proporción de C16:0 con respecto a C18:0 del total de AG saturados generalmente mejora la digestibilidad de los ácidos grasos, especialmente si el grado de insaturados de la grasa se incrementa. Zinn et al. (2000) observaron que al disminuir el índice de biohidrogenación mediante la protección de la grasa con una matriz de formaldehído-proteína mejoró la digestión intestinal de los AG totales (87,8 vs. 80,3%). Adicionalmente detectaron que por cada 1% de incremento en la proporción de C18:1 en relación al total de AG que llegan al intestino, la digestibilidad de C18:0 aumentaba en 1% (Digestibilidad de C18:0= 85,72+1,01P18:1-15,84FI; donde P18:1 es el C18:1 ingresando al intestino expresado como porcentaje del total de los AG que llegan al duodeno y FI es la cantidad diaria de AG consumidos expresados como g de AG/kg de PV, r2= 0,99). Apoyados en el concepto de que la digestión intestinal de los ácidos grasos saturados disminuye a medida que la longitud de su cadena aumenta (Steele y Moore, 1968), es razonable esperar que el ácido palmítico (C16:0) tiene condiciones más favorables para mostrar un efecto sinérgico mayor con los ácidos grasos insaturados a nivel postruminal que el ácido esteárico (C18:0). En ese sentido, Plascencia et al. (2003a) evaluaron una fuente de grasa rica en palmítico (>97%) protegida con formaldehído mezclada al 50% con grasa amarilla, la cual es rica en insaturados (>65%), añadida a un nivel de 5% de la dieta. La combinación resultó en una mejora de la digestibilidad intestinal de los ácidos grasos de 5% cuando se comparó con las fuentes de grasas originales.
Métodos de adición
Debido a sus características físicas las grasas tienden a formar una capa o cubierta en las partículas alimenticias, principalmente fibra (Devendra y Lewis, 1974). De hecho aproximadamente un 80% del total de lípidos en rumen está en forma asociada a partículas (McAllan et al., 1983) y de 40 a 75% de las bacterias está adherido a las partículas alimenticias (Owens y Goetsch, 1988). Esto hace que por sus propiedades hidrófobas las grasas por cubrimiento físico puedan ejercer un efecto inhibitorio a la acción enzimática bacteriana interfiriendo en los procesos normales de fermentación (Devendra y Lewis, 1974) y afectando, por efectos asociativos, el valor nutricional de las grasas alimenticias. En ese sentido, se han conducido una serie de estudios con la finalidad de evaluar si el método de adición de grasa puede influir en su valor nutricional. Zinn et al. (1998) evaluaron la adición de grasa amarilla, bien sea añadida primeramente al grano (maíz en hojuela) y el producto resultante mezclado con el resto de los ingredientes de la dieta, o bien la grasa amarilla era añadida al final del mezclado. En ambas situaciones la cantidad final de grasa en la dieta fue de 5%. No se detectaron diferencias entre los métodos de adición sobre los parámetros digestivos o el comportamiento productivo del ganado.
En virtud de que el punto de fusión de las grasas afecta su grado de lipólisis (Beam et al., 2000), una fuente de grasa de mayor punto de fusión podría tener un efecto diferente en cuanto a su método de adición. Para tal efecto Plascencia et al. (2001), mediante una prueba de digestión y metabolismo evaluaron, empleando el mismo esquema de tratamientos utilizados por el estudio de Zinn et al. (1998), si el método de adición puede ser afectado por el punto de fusión de la fuente de grasa utilizada (sebo vs. grasa amarilla). Al igual que el estudio de Zinn et al. (1998), no existieron efectos de los tratamientos sobre el sitio o grado de digestión de los diferentes componentes de la dieta. La digestión intestinal de la grasa promedió 69,3%. El realizar mezcla de grasa-forraje también ha sido evaluado en dietas de finalización para bovinos en engorda. La mezcla de 80% de alfalfa con 20% de grasa fue utilizada en 15, 30 y 45% en dietas de finalización. La mezcla al nivel de 15% contuvo 71% de trigo en hojuela y en los niveles de 30 y 45% fue incorporada sustituyendo directamente al trigo en hojuela. El sustituir el grano con la mezcla alfalfa-grasa no afectó el consumo o la ganancia diaria; sin embargo, la EN de la dieta disminuyó en forma lineal al aumentar la proporción de la mezcla en la dieta. De igual manera, la digestibilidad intestinal de los ácidos grasos disminuyó a medida que se aumentaba la mezcla en la ración. Se concluyó que la cantidad de grasa consumida fue el factor principal que limitó el contenido de EN y la digestibilidad de los AG, mientras que el método de adición no tuvo un papel relevante en los resultados (Plascencia y Zinn, 2002).
Con la finalidad de comprobar los resultados obtenidos en los estudios anteriores se realizó un experimento de comportamiento productivo con 216 novillos (Zinn y Plascencia, 2004a). Se evaluaron tres métodos de adición de grasa (ácidos grasos de sebo añadido bien en el grano, en el forraje o en la ración completa) con tres niveles de inclusión (3, 6 y 9%). El aumento del nivel de grasa en la dieta disminuyó el consumo, la ganancia diaria de peso y la EN de la grasa en forma lineal. Nuevamente el nivel de consumo fue el factor primordial, mientras que el método de adición no mostró efectos asociativos sobre el valor nutricional de la grasa adicionada.
Nivel de inclusión
Las recomendaciones para el uso de grasas alimenticias para dietas de rumiantes indican que éstas no deben exceder el 5% de la dieta, puesto que se han observado efectos detrimentales sobre el consumo y la eficiencia alimenticia cuando la grasa se incluye en niveles superiores (Haaland et al., 1981; Ngidi et al., 1990; Zinn, 1994). Sin embargo, las restricciones prácticas para su óptima utilización no han sido aún resueltas, ya que se han registrado casos negativos en comportamiento productivo con niveles de inclusión igual o menor al 3% (Hatch et al., 1972; Krehbiel et al., 1995), mientras que niveles de 8% han conducido a ganancias y conversiones superiores con relación a animales no suplementados (Zinn, 1989a). Lo anterior se refleja en una variabilidad del valor nutricional observado para la energía neta (EN) de la grasa que oscila desde 3,77 y 2,95 (Clary et al., 1993) hasta 6,35 y 5,15Mcal·kg-1 (Plascencia et al., 2002) de EN de mantenimiento y EN de ganancia, respectivamente, tal como se aprecia en la Tabla II.
Aún así, una respuesta generalizada cuando se aumenta el nivel de grasa en la dieta es la disminución del valor energético de la grasa. Por ejemplo, Zinn y Plascencia (2004a) informan que al aumentar el nivel de grasa de 3 a 9% en una dieta de finalización para novillos, se observaron disminuciones en el consumo, ganancia diaria y conversión alimenticia. La EN de la dieta resultó ser 103% para el nivel de inclusión de 3% y descendió a 90% cuando la grasa fue añadida en un 9% en la dieta. La EN calculada de la grasa declinó de 6,4 a 3,44Mcal·kg-1. La razón para lo anterior ha sido atribuido principalmente a variaciones en la digestión intestinal de lípidos (Wu et al., 1991; Zinn, 1994). El valor de la energía bruta (EB) para las grasas es de 9,4Kcal·g-1; sin embargo, el valor combustible "utilizable" por los tejidos debe considerarse a partir de su digestibilidad. Las grasas de grado alimenticio contienen aproximadamente 90% de ácidos grasos totales (AFOA, 1999), y éstos representan casi el 100% de su contenido energético (Zinn, 1989a). Por lo tanto, el valor energético de las grasas está supeditado a la digestibilidad de sus ácidos grasos la cual en bovinos es aproximadamente del 77% (Tabla III). Bauchart (1993) indica un 80% de digestibilidad para los AG saturados y de 93% para los insaturados cuando se consumen dietas con moderado contenido de lípidos (2-3%). La elevada capacidad de los rumiantes para digerir AG saturados, con respecto a los no rumiantes, se atribuye a las condiciones de pH duodenal (2-2,5, Christiansen y Webb, 1990), la proporción de fosfatidilcolina (80%) con alto contenido de oleico (Christie, 1973), así como por la elevada proporción de taurocolatos (Peric-Golia y Socic, 1968) de las sales biliares. Por otra parte, el concepto generalizado de que la longitud de la cadena en los AG saturados afecta negativamente a su digestibilidad (Steele y Moore, 1968) no parece ser de importancia en rumiantes cuando los consumos de lípidos son moderados (Weisjberg et al., 1992). Sin embargo, a altos consumos, la digestibilidad de C18:0 es 5 a 8% menor en relación a C16:0 (Bauchart, 1993). Como se mencionó anteriormente, la digestibilidad de los ácidos grasos insaturados es mayor que la de los saturados y no parecen ser afectados aún con altos consumos de grasa (Ávila et al., 2000; Enjalbert et al., 2000; Plascencia et al., 2003b). Existen informes que indican que el ácido linolénico es absorbido menos eficientemente que el palmítico (Doreau y Ferlay, 1994); sin embargo, esos resultados son aparentes, ya que esas diferencias se atribuyen a problemas metodológicos en la determinación de C18:3, dada su baja concentración (<2%) en el flujo duodenal.
Lo anterior demuestra, en gran medida, que el principal factor que afecta la digestibilidad de la grasa alimenticia en rumiantes es su nivel de consumo (Palmquist y Conrad, 1980; Ngidi et al., 1990; Coppock y Wilks, 1991; Palmquist, 1991; Khorasani et al., 1992; Zinn, 1992, 1994; Pylot et al., 2000). Por ejemplo, Zinn (1989b) observó una disminución en forma lineal (83, 81 y 74%) de la digestibilidad intestinal de los ácidos grasos cuando se aumentó el nivel de inclusión de grasa amarilla en la dieta de 0 a 8%, calculando una disminución de la digestibilidad de la grasa en 3,4% por cada unidad porcentual por encima de 4% de inclusión. Similares a lo anterior son los resultados de Palmquist (1991), quien observó que la digestibilidad de los ácidos grasos disminuía en 2,2% por cada 100g de ácidos grasos consumidos. Adicionalmente, Palmquist y Conrad (1980) indican que a niveles moderados de inclusión (por debajo de 5%) la digestibilidad verdadera de la grasa es aproximadamente de 80%, mas si se agrega la grasa por encima de ese nivel se digiere menos eficientemente (56%). Por otra parte, Wu et al. (1991) determinaron que la reducción de la digestibilidad total de ácidos grasos se debía principalmente al efecto negativo de la digestibilidad del ácido esteárico (C18:0), ya que registraron una disminución en su digestibilidad de un 21% cuando la grasa suplementada fue incrementada en la dieta del 3 al 6% (72 y 59%, respectivamente). Estos datos son muy similares al 75% y 59% de digestibilidad para C18:0 obtenidos por Zinn (1992) en dietas con 90% de concentrados suplementados con 0% y 6% de grasa animal.
TABLA II / COMPARACIÓN DEL CONTENIDO DE ENERGIA NETA* DE MANTENIMIENTO Y DE GANANCIA DE GRASAS ALIMENTICIAS ESTIMADO POR MEDIO DE LA TÉCNICA DE REEMPLAZO EN NOVILLOS CON DIETAS DE FINALIZACIÓN
* Mcal·kg-1.
TABLA III / DIGESTIBILIDAD INTESTINAL PARA RUMIANTES DE LAS DISTINTAS FUENTES DE GRASA
a MAV: mezcla animal-vegetal, GA: grasa amarilla, TG: triglicéridos, SV: sebo vacuno.
Los estudios anteriores muestran concordancia en que a mayor nivel de inclusión menor es la eficiencia de utilización intestinal de los ácidos grasos, principalmente los saturados (C16:0 y C18:0). Sin embargo, no existe concordancia respecto a qué porcentaje de inclusión permite el máximo de utilización de grasa sin afectar negativamente su digestibilidad, y por lo tanto su contenido de EN. Aún así, las recomendaciones prácticas actuales indican que la inclusión no exceda del 5% de la dieta. Aunque Doreau y Ferlay (1994), al relacionar la digestibilidad intestinal con el consumo de grasa de 13 experimentos, no encontraron relación entre el nivel de grasa en la dieta y la digestión de los ácidos grasos, es importante hacer notar que la relación establecida por esos investigadores fue la de la concentración de lípidos (g·kg-1 de MS) frente a digestibilidad expresada en porcentaje. Esta relación puede confundir los efectos, ya que indica sólo la concentración de lípidos en la dieta y no la cantidad total de lípidos consumidos. Respecto a lo anterior, Brandt et al. (1992) indican que mucha de la inconsistencia en la respuesta del comportamiento productivo del ganado cuando consume dietas suplementadas con grasa puede estar más asociada con la cantidad total de lípidos consumidos que con el porcentaje de inclusión.
Al evaluar el efecto del nivel de consumo de ácidos grasos sobre el valor nutricional de las grasas relacionándolo con su digestión intestinal, Plascencia et al. (2003b) observaron una correlación negativa de la cantidad de AG consumidos (g·kg-1 de peso corporal) frente a su digestibilidad intestinal. Combinando los resultados obtenidos en ese estudio con otros 7 estudios previos se generó la ecuación de regresión (Figura 1)
y= 87,56 - 8,591x (R2= 0,89, n=25)
donde y: digestibilidad intestinal, y X: ácidos grasos consumidos por día (g·kg-1 de peso corporal). Esta regresión implica que para no afectar a la digestibilidad intestinal de los AG, el consumo diario de ácidos grasos totales debe ser menor a 0,96g de AGT por kg de peso corporal.
Figura 1. Influencia del nivel de consumo de ácidos grasos (AG)sobre su digestión intestinal.
Lo anterior indica que la principal limitante en la digestibilidad sucede cuando la cantidad de los lípidos que llegan al duodeno sobrepasa la capacidad enzimática intestinal y, por consiguiente, su absorción (Bauchart, 1993). Los ácidos grasos que llegan al intestino en rumiantes son altamente saturados (>65%) y están asociados a partículas (Wu et al., 1991; Pantoja et al., 1996; Plascencia et al., 1999). Los AG saturados son físicamente menos voluminosos y más viscosos que los insaturados, lo cual dificulta la formación de la fase micelar (Moore y Christie, 1984) y la tasa de absorción, puesto que se ha comprobado que la absorción de los AG saturados es más lenta (Doreau y Ferlay, 1994). Debe considerarse que la forma de evaluar la digestibilidad se basa no sólo en el potencial de digestión del nutriente en el organismo, sino también en su potencial de absorción en el tracto gastrointestinal, y es importante que se considere esta limitante cuando se presentan altos consumos de lípidos. La disminución de la digestibilidad de los AG saturados puede explicar de 85 a 100% la variación del valor nutricional observado para las grasas adicionadas a las dietas para rumiantes (Pantoja et al., 1995; Ramírez y Zinn, 2000; Ávila et al., 2000; Enjalbert et al., 2000) y esto puede estar relacionado con una posible emulsificación incompleta de los AG saturados en el intestino delgado como resultado de una producción inadecuada de jugo biliar con relación a la cantidad presente de estos ácidos grasos que se observa como resultado del consumo de dietas con alto contenido de grasa (Palmquist, 1991; Xu et al., 1998). En ese sentido, en aves se ha demostrado deficiencias en la utilización adecuada de las grasas alimenticias por insuficiencia de jugo biliar. Atteh y Lesson (1985) observaron un incremento del 16% en la digestión intestinal de una mezcla de palmítico:oleico cuando se añadió 0,2% de ácido cólico a la dieta, lo que se tradujo en una mejora de un 14% en la energía digestible de la dieta. Considerando que en rumiantes la producción de jugo biliar oscila de 0,5 a 1,45ml·kg-1 de peso (Harrison, 1962; Moore y Christie, 1984; Merchen, 1988), para un bovino de 200kg de peso vivo la producción biliar varía desde 2400 hasta 6960ml/día. Sin embargo, las diferencias detectadas en la producción biliar evaluada por métodos de cateterización del conducto biliar utilizando técnicas de recolecciones repetidas de muestras, pueden ser debidas a la disminución de la resorción de jugos biliares en las partes distales del intestino (reflejo enterocólico; Hurwitz et al., 1973) que afectan la producción biliar a corto plazo, a la disminución del flujo biliar debido a irritación directa en el conducto biliar (Harrison, 1995), a la inexactitud en la técnica de recolección de la totalidad jugo biliar (Merchen, 1988), o bien a características propias de la dieta. Con respecto a este último punto, aún cuando se ha determinado que el flujo biliar en rumiantes tiende a ser independiente de la hora del día o la frecuencia alimenticia (Symonds et al., 1982), es importante considerar a la cantidad de ácidos grasos y a la proporción de insaturados:saturados que ingresan al duodeno como una variable potencial sobre la producción biliar, ya que éstos tienen un efecto directo sobre la producción de gastrina y colecistoquinina, ambos elementos involucrados en el mecanismo de flujo biliar al duodeno (Studzinsky y Bobowiec, 1979; Kato et al., 1991).
TABLA IV / PRODUCCIÓN BILIAR Y DIGESTIÓN POSRUMINAL DE LOS ÁCIDOS GRASOSTOTALES EN NOVILLOS* CONSUMIENDO DIETAS CON 3 NIVELES DEGRASA SUPLEMENTARIA
* De 215kg de peso.
a Efecto lineal del nivel de grasa, P<0,01.
b Efecto lineal del nivel de grasa, P<0,10.
Se ha argumentado que el grado de formación de micelas, primer paso para la absorción de lípidos, puede variar con el tipo de dieta, lo que podría ocurrir sólo si se aumenta la producción biliar o la proporción de sales biliares:lípidos, o bien si la proporción de ácidos grasos insaturados:saturados en el duodeno aumenta. Sin embargo, existen estudios en los que se ha probado el uso de emulsificantes, principalmente lecitina, combinados con la grasa alimenticia en los cuales no se han obtenido beneficios en la digestión postruminal de los ácidos grasos (Zinn, 1989b; Jenkins y Fotohui, 1990; Plascencia et al., 1991). Lo anterior puede explicarse por la alta hidrólisis ruminal de la lecitina, lo que evita que llegue con sus características emulsificantes al intestino (Jenkins, 1990). Un indicio de la limitada capacidad de emulsificación a nivel intestinal en rumiantes lo muestra el estudio de Børsting et al. (1992), quienes indican coeficientes de digestión de 92% para C18:0 cuando las vacas consumieron dietas con grasas vegetales emulsificadas y protegidas contra la hidrólisis ruminal mediante una matriz de formaldehído-caseína. Esos investigadores concluyeron que los bajos coeficientes de digestión observados en la mayoría de los estudios para C18:0 con altos consumos de lípidos son debidos principalmente a una pobre emulsificación más que a un inefectivo sistema de lipasas. En ese sentido, Plascencia et al. (2004) relacionaron la producción biliar con la digestión postruminal de los AG en novillos que consumieron dietas con tres distintos niveles de grasa amarilla. La producción biliar promedió 32ml·kg-1 de PV y se incrementó (efecto lineal) a medida que se aumentó el flujo de lípidos al duodeno. Sin embargo, considerada como proporción del flujo de lípidos (ml de bilis por g de lípido que ingresa al duodeno proximal) la producción biliar disminuyó linealmente de 23,4 a 16,7ml de bilis/g de lipidos en duodeno (Tabla IV). La mayor digestibilidad para AG se observó en una relación de 40ml de bilis/ g de lípidos en duodeno y la producción biliar explicó en 69% la variación en la utilización de AG en ese experimento (Figura 2).
Figura 2. Influencia de la producción biliar (ml de bilis por g delípidos en duodeno) sobre la digestión intestinal de los AG.
Conclusiones
Independientemente de la fuente y la forma de adición de grasa a la dieta, el valor nutricional de las grasas asignados por el NRC (1996) tiende a ser consistente cuando el consumo total de grasas está dentro de la proporción de 0,86 a 0,96g de grasa consumida/kg peso vivo. Cuando el consumo de grasa es mayor a esa proporción, el valor energético de la grasa disminuye en forma lineal como resultado directo de la disminución de la digestibilidad intestinal de los ácidos grasos, principalmente C18:0, posiblemente por una limitada capacidad de producción biliar. La digestibilidad intestinal de los ácidos grasos es una función altamente predecible a partir de su consumo total por unidad de peso corporal.
REFERENCIAS
1. AFOA (1999) Trading and Arbitration Rules. American Fats and Oils Association. New York, EEUU. pp. 34-36.2. Atteh JO, Lesson S (1985) Influence of age dietary cholic acid and calcium levels on performance, utilization of free fatty acids and bone mineralization in broilers. Poultry Sci. 64: 1959-1971.3. Ávila CD, DePeters EJ, Pérez-Monti H, Taylor SJ, Zinn RA (2000) Influences of saturation ratio of supplemental dietary fat on digestion and milk yield in dairy cows. J. Dairy Sci. 83: 1505-1519.4. Barreras-Arellano D (1998) Estabilidad y utilización de nitrógeno en grasas y aceites. Instituto de la grasa, CSIC, España. Rev. Grasas y Aceites. 49: 56-64.5. Bauchart D (1993) Lipid absorption and transport in ruminants. J. Dairy Sci. 76: 3864-3881.6. Beam TM, Jenkins TC, Moate PJ, Khon RA, Palmquist DL (2000) Effects of amount and source of fat on the rates of lipolysis and biohydrogenation of fatty acids in ruminal contents. J. Dairy Sci. 83: 2564-2573.7. Bock BJ, Harmon DL, Brandt RTJr, Schneider JE (1991) Fat source and calcium level effects on finishing steer performance, digestion, and metabolism. J. Anim. Sci. 69: 2211-2224.8. Børsting CF, Weisjberg MR, Hvelplund TH (1992) Fatty acid digestibility in lactating cows fed increasing amounts of protected vegetable oil, fish oil or saturated fat. Acta. Agric. Scand. A. Animal Sci. 42: 148-156.9. Brandt RTJr, Anderson SJ (1990) Supplemental fat source affects feedlot performance and carcass traits of finishing yearling steers and estimated diet net energy value. J. Anim. Sci. 68: 2208-2216.10. Brandt RTJr, Kuhl GL, Campbell RE, Kastner CL, Stroda SL (1992) Effects of steam-flaked sorghum grain or corn and supplemental fat on feedlot performance, carcass traits, longissimus composition, and sensory properties of steers. J. Anim Sci. 70: 343-348.11. Byers FM, Shelling GT (1988) Lipids in ruminant nutrition. En Church DC (Ed.) The ruminant animal: Digestive physiology and nutrition. Prentice-Hall. New Jersey, EEUU. pp. 289-312.12. Cera KR, Mahan DC, Reinhart GA (1989) Apparent fat digestibilities and performance response of postweaning swine fed diets supplemented with coconut oil, corn oil, or tallow. J. Anim. Sci. 67: 2040-2047.13. Christie WW (1973) The structures of bile phosphatidylcholines. Biochim. Biophys. Acta. 316: 204-211.14. Christiansen ML, Webb KEJr (1990) Intestinal acid flow, dry matter, starch and protein digestibility and amino acid absorption in beef cattle fed a high concentrate diet with defluorinated rock phosphate, limestone or magnesium oxide. J. Anim. Sci. 68: 2105-2118.15. Clary EM, Brandt TRJr, Harmon DL, Nagaraja TG (1993) Supplemental fat and ionophores in finishing diets: Feedlot performance and ruminal digesta kinetics. J. Anim. Sci. 71: 3115-3123.16. Coppock CE, Wilks DL (1991) Supplemental fat in high-energy rations for lactating cows: Effects on intake, digestion, milk yield, and composition. J. Anim. Sci. 69: 3826-3837.17. Czerkawsky JW (1973) Effect of linseed oil fatty acids and linseed oil rumen fermentation in sheep. J. Agric. Sci. 81: 517-531.18. Demeyer DI, Hederickx HK (1967) The effect of C18 unsaturated fatty acids on methane production in vitro by mixed rumen bacteria. Biochim. Biophys. Acta. 137: 484-497.19. Devendra CA, Lewis D (1974) Interaction between dietary lipids and fiber in the sheep. Anim. Prod. 19: 67-76.20. Doreau M, Ferlay A (1994) Digestion and utilization of fatty acids by ruminants. Anim. Feed Sci. Technol. 45: 379-396.21. Doreau M, Chilliard Y, Rulquin H, Demeyer DI (1999) Manipulation of milk fat in dairy cows. En Garnsworthy PC, Wiseman J (Eds.) Recent Advances in Animal Nutrition. Nottingham University Press. Nottingham, RU. pp. 81-110.22. Duckett SK, Andrae JG, Owens FN (2002) Effect of high-oil corn or added corn oil on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finishing diets. J. Anim. Sci. 80: 3353-3360.23. Elizalde JC, Aldrich CG, LaCount DW, Drackley JK, Merchen NR (1999) Ruminal and total tract digestibilities in steers fed diet containing liquefied or prilled saturated fatty acids. J. Anim. Sci. 77: 1930-1939.24. Elliott JP, Drakley JK, Aldrich CG, Merchen NR (1997) Effect of saturation and esterification of fat sources on site and digestion of organic matter, fiber and nitrogen. J. Anim. Sci. 75: 2803-2812.25. Elliott JP, Drackley JK, Beaulieu AD, Aldrich CG, Merchen NR (1999) Effect of saturation and esterification of fat sources on site and extent of digestion in steers: Digestion of fatty acids, triglycerides, and energy. J. Anim. Sci. 77: 1919-1929.26. Enjalbert F, Nicot MC, Bayourthe C, Moncoulon R (2000) Effect of duodenal infusion of palmitic, stearic or oleic acids on milk composition and physical properties of butter. J. Dairy Sci. 83: 1428-1433.27. Firkins JL, Eastridge ML (1994) Assesment of the effect of iodine value on fatty acid digestibility, feed intake and milk production. J. Dairy Sci. 77: 2357-2366.28. Garnsworthy PC (2002) Fats in dairy cow diets. En Garnsworthy PC, Wiseman J (Eds.) Recent Advances in Animal Nutrition 4. Nottingham University Press. Hampshire, Inglaterra. pp. 399-416.29. Haaland GL, Matsushima JK, Jhonson DE, Ward GM (1981) Effect of replacement of corn by protected tallow in a cattle finishing diet on animal performance and composition. J. Anim. Sci. 52: 696-702.30. Hamilton CR (2002) Value of animal fats and recycled greases in animal feeds. Darling International Inc. Irving, TX, EEUU. 17 pp.31. Harfoot CG, Hazlewood GP (1997) Lipid metabolism in the rumen. En Hobson PN (Ed.) The rumen microbial ecosystem. Elsevier. London, New York. pp 382-426.32. Harrison FA (1962) Bile secretion in the sheep. J. Physiol. 162: 212-224.33. Harrison FA (1995) Surgical techniques in experimental farm animals. Oxford University Press. Oxford, RU. pp. 174-186.34. Hatch CF, Perry TW, Mohler MT, Beeson WM (1972) Effect of added fat with graded levels of calcium and urea-containing rations for beef cattle. J. Anim. Sci. 34: 483-487.35. Hawke JC, Silcock WR (1970) The in vitro rate of lipolysis and biohydrogenation in rumen contents. Biochim. Biophys. Acta 218: 201-212.36. Henderson C (1973) The effects of fatty acids on pure cultures of rumen bacteria. J. Agric. Sci. 81: 107-112.37. Huffman RP, Stock RA, Sindt MH, Shain DH (1992) Effect of fat type and forage level on performance of finishing cattle. J. Anim. Sci. 70: 3889-3898.38. Hurwitz S, Bar A, Katz M, Skan D, Budowsky P (1973) Absorption and secretion of fatty acids and bile acids in intestine of the laying fowl. J. Nutr. 103: 543-547.39. Inming I, Van Nevel C, Demeyer DI (1993) Lipolysis and hydrogenation of soybean oil in the rumen of sheep. Proc. Soc. Nutr. Physiol. Willingeen, Alemania. pp. 59-64.40. Jenkins TC (1990) Nutrient digestion, ruminal fermentation and plasma lipids in steers fed combinations of hydrogenated fat and lecithin. J. Dairy Sci. 73: 2934-2939.41. Jenkins TC (1993) Lipid metabolism in the rumen. J. Dairy Sci. 76: 3851-3863.42. Jenkins TC, Fotohui N (1990) Effects of lecithin and corn oil on site of digestion, ruminal fermentation and microbial protein synthesis. J. Anim. Sci. 68: 460-466.43. Jenkins TC, Jenny BL (1989) Effect of hydrogenated fat on feed intake, nutrient digestion and lactational performance of dairy cows. J. Dairy Sci. 72: 2316-2324.44. Johnson RR, McClure KE (1972) High fat rations for ruminants. I. The addition of saturated and unsaturated fats to high roughage and high concentrate rations. J. Anim. Sci. 34: 501-509.45. Kato S, Katoh K, Barej W (1991) Regulation of exocrine pancreatic secretion in ruminants. En Tsuda T, Sasaki Y, Kawashima R (Eds.) Physiological aspects of digestion and metabolism in Ruminants. Proc. 7th Int. Symp. Ruminal Physiol. Academic Press. San Diego, CA, EEUU. pp. 89-110.46. Khorasani GR, de Boer G, Robinson PH, Kennedy JJ (1992) Effect of canola fat on ruminal and total tract digestion, plasma hormones and metabolites in lactating dairy cows. J. Dairy Sci. 75: 492-501.47. Klusmeyer TH, Clark JH (1991) Effects of dietary fat and protein on fatty acids flow to the duodenum and milk produced by dairy cows. J. Dairy Sci. 74: 3055-3067.48. Krehbiel CR, McCoy RA, Stock RA, Klopfenstein TJ, Shain DH, Huffman RP (1995) Influence of grain type, tallow level, and tallow feeding system on feedlot cattle performance. J. Anim. Sci. 73: 2916-2921.49. Latham MJ, Storry JE, Sharpe ME (1972) Effect of low-roughage diet on the microflora and lipid metabolism in the rumen. Appl. Microbiol. 24: 871-877.50. Lofgreen GP (1965) Net energy of fat and molasses for beef heifers with observations on method for net energy determination. J. Anim. Sci. 24: 480-487.51. MacLeod GK, Buchannan-Smith JG (1972) Digestibility of hidrogenated tallow, saturated fatty acids and soybean oil supplemented diets by sheep. J. Anim. Sci. 35: 890-895.52. Maczulak AE, Dehority BA, Palmquist DL (1981) Effects of long-chain fatty acids on growth of rumen bacteria. Appl. Envir. Microbiol. 42: 856-862.53. McAllan AB, Knight BR, Sutton JD (1983) The effect of free and protected oils on the digestion of dietary carbohydrates between the mouth and duodenum of sheep. Br. J. Nutr. 48: 433-440.54. Merchen NR (1988) Digestion, absorption and excretion in ruminants. En Church DC (Ed.) The ruminant animal: Digestive physiology and nutrition. Prentice-Hall. New Jersey, EEUU. pp. 172-201.55. Moore JH, Christie WW (1984) Digestion, absorption and transport of animal fats in ruminant animal. En Wiseman J (Ed.) Fats in Animal Nutrition. Butterworths. Londres, Inglaterra. pp. 123-149.56. Moore JA, Swingle RS, Hale WH (1986) Effects of whole cottonseed, cottonseed oil or animal fat on digestibility of wheat straw diets by steers. J. Anim. Sci. 63: 1267-1273.57. Ngidi ME, Loerch SC, Fluharty FL, Palmquist DL (1990) Effect of calcium soaps of long-chain fatty acids on feedlot performance carcass characteristics and ruminal metabolism in steers. J. Anim. Sci. 68: 2555-2565.58. Noble RC, Moore JH, Harfoot CG (1974) Observations of the pattern of biohydrogenation of sterified and unsterified linoleic acid in the rumen. Br. J. Nutr. 31: 99-108.59. NRC (1996) Nutrient Requirements of Beef Cattle. 7th ed. National Academy of Sciences Press. Washington, DC, EEUU. pp. 133-146.60. Owens FN, Goetsch AL (1988) Ruminal fermentation. En Church DC (Ed.) The Ruminant Animal: Digestive Physiology and Nutrition. Prentice-Hall, New Jersey, EEUU. pp. 145-171.61. Palmquist DL (1991) Influence of source and amount of dietary fat on digestibility in lactating cows. J. Dairy Sci. 74: 1354-1360.62. Palmquist DL, Conrad HR (1980) High fat rations for dairy cows. Tallow and hydrolyzed blended fat at two intakes. J. Dairy Sci. 63: 391-395.63. Palmquist DL, Weisjberg MR, Hvelplund T (1993) Ruminal, intestinal, and total tract digestibilities of nutrients in cows fed diets high in fat and undegradable protein. J. Dairy Sci. 76: 1356-1364.64. Pantoja J, Firkins JL, Eastridge ML (1995) Site of digestion and milk production by cows fed fats differing in saturation, esterification, and chain length. J. Dairy Sci. 78: 2247-2258.65. Pantoja J, Firkins JL, Eastridge ML (1996) Fatty acid digestibility and lactation performance by dairy cows fed fats varying in degree of saturation. J. Dairy Sci. 79: 429-437.66. Peric-Golia L, Socic C (1968) Biliary bile acids and cholesterol in developing sheep. Am. J. Physiol. 215: 1284-1287.67. Plascencia A, Zinn RA (2001) Comparative feeding value of tallow vs yellow grease in finishing diets for feedlot cattle. Proc. West. Sect. Am. Soc. Anim. Sci. 52: 566-568.68. Plascencia A, Zinn RA (2002) Evaluation of a forage-fat blend as an isocaloric substitute for steam-flaked wheat in finishing diets for feedlot cattle: Growth-performance and digestive function. Prof. Anim. Sci. 18: 247-253.69. Plascencia A, Álvarez EG, Zinn RA (1991) Efecto de lecitina y grasa suplementaria sobre digestión de nutrientes y fermentación ruminal en dietas para cabras lactantes. Rev. Cs. Agropec. 3: 49-58.70. Plascencia A, Estrada M, Zinn RA (1999) Influence of free fatty acid content on the feeding value of yellow grease in finishing diets for feedlot cattle. J. Anim. Sci. 77: 2603-2609.71. Plascencia A, Cervantes MA, Zinn RA (2001) Influence of fat titer and method of addition on characteristics of ruminal and total tract digestion. Proc. West. Sect. Am. Soc. Anim. Sci. 52: 548-550.72. Plascencia A, Álvarez EG, Montano MF, Machado M, Rodríguez S, Ware RA, Zinn RA (2002) Influence of level of intake on the comparative feeding value of fat in finishing diets for feedlot cattle. Proc. West. Sect. Am. Soc. Anim. Sci. 53: 613-618.73. Plascencia A, Corona-Gochi L, Ware RA, Zinn RA (2003a) Influencia de la proporción entre ácidos grasos insaturados y ácido palmítico que fluyen a duodeno sobre la digestión intestinal de ácidos grasos en novillos con dietas de crecimiento-finalización suplementadas con grasa. XIII Reunión Internacional de Producción de Carne y Leche en Climas Cálidos. Mexicali, México. pp. 185-189.74. Plascencia A, Vázquez C, Mendoza GD, Zinn RA (2003b) Relationship between body weight and level of fat supplementation on fatty acid digestion in feedlot cattle. J. Anim. Sci. 81: 2653-2659.75. Plascencia A, Vázquez C, Mendoza GD, Zinn RA (2004) Influences of levels of fat supplementation on bile flow and fatty acid digestión in cattle. J. Anim. Vet. Adv. 11: 763-768.76. Polan CC, McNeill JJ, Store SB (1964) Biohydrogenation of insaturated fatty acid by rumen bacteria. J. Bacteriol. 88: 1056-1064.77. Pylot SJ, McKinnon JJ, Mustafa AF, Racz VJ, Christensen DA (2000) Effects of processing and fat content of coarse canola screenings on voluntary intake and total tract nutrient digestibility of beef steers. Can. J. Anim. Sci. 80: 153-159.78. Ramírez JE, Zinn RA (2000) Interaction of dietary magnesium level on the feeding value of supplemental fat in finishing diets for feedlot steers. J. Anim. Sci. 78: 2072-2080.79. Robert WK, McKirdy JA (1964) Weight gains, carcass fat characteristics and ration digestibility in steers as affected by dietary rapeseed oil, sunflowerseed oil, and animal tallow. J. Anim. Sci. 63: 682-687.80. Steele W, Moore JH (1968) The digestibility coefficients of myristic, palmitic and stearic acids in the dit of sheep. J. Dairy Res. 35: 371-376.81. Studzinski T, Bobowiec R (1979) Effect of intra-duodenal administration of HCl and NaHCO3 on the secretion and content of sodium, potassium and calcium in the pancreatic juice and bile of sheep. Pol. Arch. Weter. 22: 205-217.82. Symonds HW, Mather DL, Hall ED (1982) Surgical procedure for modifying the duodenum in cattle to measure bile flow and the diurnal variation in biliary manganese, iron, copper and zinc excretion. Res. Vet. Sci. 32: 6-11.83. Valenzuela AB (1995) Natural antioxidants: A new perspective for the problems of oxidative rancidity of lipids. En Lyons TP, Jacques KA (Eds.) Biotechnology in the Feed Industry. Proc. 11th Ann. Symp. Nottingham University Press, Loughborough, Leicester, RU. 207-220 pp.84. Van Nevel CJ, Demeyer DI (1996) Influences of pH on lipolysis and biohydrogenation of soybean oil by rumen contents in vitro. Reprod. Nutr. Dev. 36: 53-63.85. Vila B, Esteve-García E (1996) Studies on acid oils and fatty acids for chickens. II. Effect of free fatty acid content and degree of saturation of free fatty acids and neutral fat on fatty acid digestibility. Br. Poultry Sci. 37: 119-130.86. Wachira AM, Sinclair LA, Wilkinson RG, Hallett K, Enser M, Wood JD (2000) Rumen biohydrogenation of n-3 polyunsaturated fatty acids and their effects on microbial efficiency and nutrient digestibility in sheep. J. Agric. Sci. 135: 419–428.87. Weisjberg MR, Hvelplund T, Børsting CF (1992) Digestibility of fatty acids in gastrointestinal tract of dairy cows fed with tallow or saturated fats rich in stearic acid or palmitic acid. Acta Agric. Scand. A. Anim. Sci. 42: 115-120.88. Wu Z, Ohajuruka OA, Palmquist DL (1991) Ruminal synthesis, biohydrogenation, and digestibility of fatty acids by dairy cows. J. Dairy Sci. 74: 3025-3034.89. Wu Z, Huber JT, Sleiman FT, Simas JM, Chen KH, Chan SC, Fontes C (1993) Effect of three supplemental fat sources on lactation and digestion in dairy cows. J. Dairy Sci. 76: 3562-3570.90. Xu C, Wensing T, Beynen AC (1998) Effects of high calcium intake on fat digestion and bile acid excretion in feces of veal calves. J. Dairy Sci. 81: 2173-2177.91. Yokoyama MT, Johnson KA (1988) Microbiology of the rumen and intestine. En Church DC (Ed.) The ruminant animal: Digestive physiology and nutrition. Prentice-Hall, New Jersey, EEUU. pp. 125-144.92. Zinn RA (1988) Comparative feeding value of supplemental fat in finishing diets for feedlot steers supplemented with and without monensin. J. Anim. Sci. 66: 213-227.93. Zinn RA (1989a) Influence of level and source of dietary fat on its comparative feeding value in finishing diets for steers: Feedlot cattle growth performance J. Anim. Sci. 67: 1029-1037.94. Zinn RA (1989b) Influence of level and source of dietary fat on its comparative feeding value in finishing diets for steers: Metabolism. J. Anim. Sci. 67: 1038-1049.95. Zinn RA (1992) Comparative feeding value of supplemental fat in steam-flaked corn- and steam-flaked wheat- based finishing diets for feedlot cattle. J. Anim. Sci. 70: 2959-2969.96. Zinn RA (1994) Effects of excessive supplemental fat on feedlot cattle growth performance and digestive function. Prof. Anim. Sci. 10: 66-72.97. Zinn RA, Álvarez EG, Plascencia A, Shen Y (1998) Influence of method of supplementation on the utilization of supplemental fat by feedlot steers. Proc. West. Sect. Am. Soc.Anim. Sci. 49: 291-296.98. Zinn RA, Plascencia A (1992) Comparative digestion of yellow grease and calcium Soaps of long-chain fatty acids in cattle. Proc. West. Sect. Am. Soc. Anim. Sci. 43: 454-457.99. Zinn RA, Plascencia A (1993) Interaction of whole cottonseed and supplemental fat on digestive function in cattle. J. Anim. Sci.. 71: 11-17.100. Zinn RA, Plascencia A (1996) Effect of forage level on the comparative feeding value of supplemental fat in growing-finishing diets. J. Anim. Sci. 74: 1194-1201.101. Zinn RA, Plascencia A (2004a) Influence of level and method of supplementation on the utilization of supplemental fat by feedlot steers. J. Anim. Vet. Adv. 3: 473-477.102. Zinn RA, Plascencia A (2004b) Future of tallow as an ingredient in livestock diets. 24th Western Nutrition Conference. Saskatoon, Saskatchewan, Canadá. pp. 21-27.103. Zinn RA, Shen Y (1996) Interaction of dietary calcium and supplemental fat on digestive function and growth performance in feedlot steers. J. Anim. Sci. 74: 2303-2309.104. Zinn RA, Gulati SK, Plascencia A, Salinas J (2000) Influence of ruminal biohydrogenation on the feeding value of fat in finishing diets for feedlot cattle. J. Anim. Sci. 78: 1738-1746.Alejandro Plascencia Jorquera. Medico Veterinario Zootecnista, Maestro y Doctor en Ciencias en Producción y Salud Animal, Universidad Nacional Autónoma de México (UNAM). Profesor Investigador, Universidad Autónoma de Baja California, Mexicali, México. Dirección: Instituto de Investigaciones en Ciencias Veterinarias, Fracc. Campestre S/N, Mexicali, Baja California, México.Germán David Mendoza Martínez. Doctor en Ciencias en Nutrición, Nebraska University, EEUU. Profesor Investigador, Colegio de Posgraduados, Montecillo, México.Carlos Vásquez Peláez. Doctor en Ciencias en Genética y Estadística, Purdue University, EEUU. Profesor Investigador, UNAM, México.Richard Avery Zinn. BS, MSc, PhD en Nutrición Animal. Profesor, University of California, El Centro, EEUU.
Temas relacionados:
Autores:

Universidad Autónoma de Baja California (UABC)
Recomendar
Comentar
Compartir

Universidad Autónoma de Baja California (UABC)
19 de abril de 2012
Estimado Miguel
Teóricamente la pre-emulsificación ayudaría a disminuir la tasa de biohidrogenación formándose menos C18:0 que es el principal factor que disminuye la digestibilidad total de ácidos grasos a nivel intestinal. Por ello se ha probado pre-emulsificar las grasas con lecitina (Jenkins y Fotohui, 1990; Plascencia et al., 1992) y con surfactantes tales como el Tween 80 (Davila-Ramos et al. 2010, 2011) sin resultados positivos. El uso de bilis es interesante pero fuera del organismo se oxida rapidamente y pierde su potencial. El uso de fosfatidilcolina es promisorio pero requiere altas concentraciones lo cual lo hace economicamente inviable. Aparentemente, en los estudios que hemos realizado no se ha usado la concentracion adecuada; sin embargo, mayores concentraciones serian muy costosas. La cantidad de grasa amarilla utilizada en concentrado de terneras es de 4%
Gracias por su interes
Recomendar
Responder
Universidad Autónoma de Baja California (UABC)
22 de enero de 2012
Estimado Miguel
Interesantisimas preguntas!
Una condición para que las acidos grasos sean saturados en el rumen es que las grasas o lípidos deben ser hidrolizados previamente mediante las esterarasas microbianas. Si hay mayor contenido de ácidos grasos libres en las grasas suplementarias la tasa de biohidrogenación ruminal disminuye y por lo tanto llegan a intestino una mayor cantidad de AG insaturados lo que los hace mas absorbibles sin embargo los AG libres tienden a afectar más la fermentación ruminal ya que son tóxicos para la microbiota. La bilis es el emulsificante natural por excelencia sin embargo fuera del organismo tiende a oxidarse fácilmente y perder su potencial emulsificador (Yo he intentado sin éxito!!). Por otra parte el uso de otros emulsificantes como lecitinas no ha tenido exito (Jenkins y Fotohui, 1990. J.Anim.Sci. 68:460-466) y nosotros probamos el uso de Tween 80 un poderosos emulsificante añadido a la grasa y no observamos benáficios sobre el valor energético de la grasa (Davila-Ramos et al., 2011. J. Applied Anim. Res.39:192-195).
En becerros prerumiantes se recomiendan grasa preemulsificadas o altas en AG insaturados ya que si no esa si pueden provocar diarreas.
Garcias por sus interesanters preguntas, saludos cordiales
Alejandro Plascencia
Recomendar
Responder

Universidad Autónoma de Baja California (UABC)
29 de diciembre de 2011
Estimado Gilberto
El coeficiente 0.96 considera los lípidos totales (tanto los suplementados como los contenidos en los ingredientes) esto es de mayor importancia cuando utilizamos en las dietas de finalización ingredientes con alto contenido en lípidos, por ejemplo, semilla de algodón, DDGS, etc. Los forrajes, como tal, aportan muy poca cantidad de lípidos (2-3%) y una parte de estos lípidos son ceras o parafinas que no tienen valor nutrimental, adicionalmente , la inclusión de forrajes es limitada en las dietas de finalización (10-15%).
Los lípidos contienen aproximadamente 10% de glicerol, este al ser liberado en rumen por las estearasas microbianas se puede transformar en propionato que al ser absorvido aporta energía. Sin embargo, la relación del valor alimenticio de las grasa con respecto a la absorción de los AG a nivel intestinal es de aproximadamente 98%
Gracias por tus interesantes preguntas!
Recomendar
Responder
Universidad Autónoma de Baja California (UABC)
5 de diciembre de 2011
Estimado Jose
El precio varia de acuerdo a la fuente de grasa (sebo, grasa amarilla, etc) y a la region
En el norte de Mexico el precio actual oscila entre los 700 a 850 dolares la tonelada
Gracias por tu pregunta
Recomendar
Responder
Universidad Autónoma de Baja California (UABC)
6 de junio de 2011
Esimado Rafael
Las sugerencias contenidas en el articulo son especificas para bovinos en engorde. Los aves como los cerdos no tienen las caracteristicas de modificar la saturación de los AG antes de que estos lleguen a intestino, por lo tanto, el grado de saturación de las grasas suplementarias para estas especies no-rumiantes si afecta negativamente la digestión y utilización de los AG y por lo tanto su valor nutrimental.
Reciba saludos cordiales
Recomendar
Responder


IDAL
27 de mayo de 2007
Muy interesante. Deseo aportar que de las grasas pasantes pueden distinguirse por lo menos tres tradicionales: los ácidos grasos fraccionados de alto punto de fusión (ac. esteárico, palmítico); las grasas o aceites hidrogenados artificialmente, y finalmente los jabones cálcicos. Existe una muy frecuente y riesgosa confusión entre los profesionales nutricionistas respecto de los jabones sódicos y los cálcicos. Los Jabones sódicos (tipo de lavar) son solubles en agua y los cálcicos (sales cálcicas de ácidos grasos) en cambio, no lo son y, por ende, sólo éstas últimas son las de considerar rumen pasantes. Las grasas protegidas con matriz proteica modificada, como un cuarto tipo de grasa pasante, son por ahora de relación costo/beneficio fuertemente negativo. Al utilizar grasas en bovinos se debe recordar en términos simples que se está quitando energía disponible a la microflora ruminal, y si no se toman las debidas precauciones podemos esperar una baja en el contenido de proteína de la leche. En general se puede recomendar que se haga un balance energético/proteico ruminal y uno pasante, es decir, al adicionar grasas pasantes acompañarlas también con un incremento de la proteína pasante.
Recomendar
Responder
Universidad Autónoma de Baja California (UABC)
13 de febrero de 2007
Estimado Oscar, la mejor respuesta del ganado cuando se incluyen grasas a las dietas, se observa cuando se da una adaptación gradual, misma que se consigue, tal como lo comentas, agregando cantidades moderadas de grasa en las dietas iniciales. Ahora bien, para tomar la decisión de la cantidad máxima de grasa que debe ser adicionada cuando los animales ya están adaptados, es importante considerar no sólo la cantidad de grasa adicionada sino el aporte total de grasa de la ración.
Recomendar
Responder

Universidad Autónoma de Baja California (UABC)
7 de febrero de 2007
Muchas gracias por tus comentarios, Diego. Y ya que tocaste el tema de grasas de sobrepaso o grasas inertes, aún cuando este tipo de grasas son muy utilizadas en ganado lechero en producción, son poco utilizadas para bovinos de engorda en confinamiento. Lo anterior obedece a que no han tenido los resultados esperados en comportamiento (Ngidi et al., 1991), y en cuanto a la protección del proceso de biohidrogenación (Zinn y Plascencia, 1992). Aún así , recientemente las grasas se han combinado con una matríz de proteína modificada (con formaldehido), que aparentemente le brinda una mejor protección en medios ruminales más ácidos (Zinn et al., 2000). Sin embargo, dado su costo su uso en las engordas, no es muy extendido.
Recomendar
Responder

30 de septiembre de 2012
Favor requiero información de microorganismos lipolíticos, actividad de las lipasas y rancidez del chocolate
Recomendar
Responder

19 de abril de 2012
Senior Alejandro:
Tenemos un ordenia ( cerca de Veracruz, Ver.), holandocebu de 20 vacas con una produccion de 8.2 lt/dia y las suplemento con 1 kg. Por vaca De concentrado al 18 % al cual le agregamos 75 gr. de Harina de carne + 50 gr. de Sal mineral. a la hora de la ordenia en la manana, despues salen a pastar Insurgentes hasta las 2 de tarde hora de amamantar los becerros. En la tarde se les sire 5 kg. silo de kingrass y se quedan en potreros de estrella hasta la maniana siguiente para su ordenia, Pregunta: Esta bien Los 75 gr. De harina de carne que les servimos o le agrego adicionalmente 25 gr. de grasa y cebo de carne en polvo? con la finalized de mejorar la condition corporal, la produccion de leche y mejorar la fertilidad.
Cabe mencionar que las vacas estan en condicion corporal regular ( casi en 3 ), y se estan preniando en promedio de 100 a 120 dais de posparto, se puede mejorar con la Grasa De Sobrepaso?.
Gracias por su respuesta y reciba un saludo desde el Tropico Humedo Caluroso de Veracruz
Recomendar
Responder


¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



.jpg&w=3840&q=75)