Efecto del amamantamiento restringido y la complementación alimenticia sobre las ganancias de peso y anestro posparto en vacas cárnicas
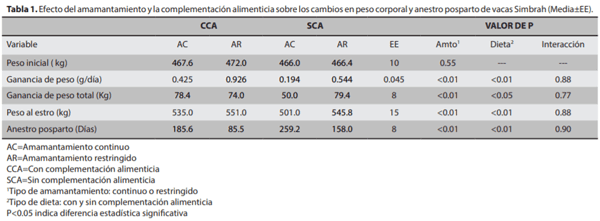
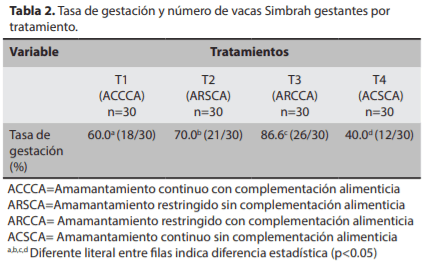
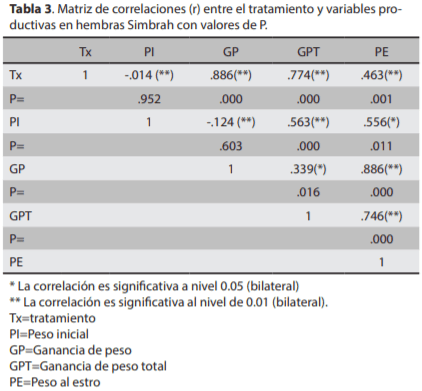
El objetivo del presente estudio fue evaluar el efecto del amamantamiento restringido y la complementación alimenticia sobre las ganancias de peso y anestro posparto, en bovinos de carne, para lo cual se utilizaron 120 vacas Sim-brah lactantes, distribuidas en cuatro tratamientos: T1) ama-mantamiento continuo con complementación alimenticia (ACCCA; n=30), T2) amamantamiento restringido sin comple-mentación alimenticia (ARSCA; n=30), T3) amamantamiento restringido con complementación alimenticia (ARCCA, n=30) y T4) amamantamiento continuo sin complementación alimenticia (ACSCA, n=30). Los resultados fueron evaluados mediante análisis de varianza en un arreglo factorial 2x2, Chi- cuadrada y correlación simple. La ganancia de peso (g/ día) y peso al estro (kg) en las vacas del T3 fue mayor (0.926 y 551.0) comparado con T1 (0.425 y 535.0), T2 (0.544 y 545.8) y T4 (0.194 y 501.0), respectivamente (P<0.01). El anestro pos-parto (días) fue menor en T3 (85.5) comparado con T1(185.6), T2 (158.0) y T4 (259.2) (P<0.01). La tasa de gestación (%) fue mayor en T3 (86.6, 26/30) con respecto a T1 (60.0, 18/30), T2 (70.0, 21/30) y T4 (40.0, 12/30) (P<0.05). Se encontró efecto por tipo de amamantamiento y CA. En conclusión, el ARCCA eleva la tasa de preñez y disminuye el anestro posparto en vacas Simbrah en el trópico.
Palabras clave: Días abiertos, nutrición, ovulación, amaman-tamiento restringido.
Arthington, J.D., Kalmbacher, R.S. 2003. Effect of early weaning on the performance of three-year-old, first-calf beef heifers and calves reared in the subtropics. Journal of Animal Science 81:1136-1141.
Arthington, J.D., Minton, J.E. 2004. The effect of early weaning on feed intake, growth, and postpartum interval in thin, Brahman-crossbred primiparous cows. The Professional Animal Scientist 20:34-38.
Barb, C.R., Kraeling, R.R. 2004. Role of leptin in the regulation of gonadotropin secretion in farm animals. Animal Reproduction Science 82-83: 155-167.
Barreiros, T.R.R., Blaschi, W., Santos, G.M.G., Morotti, F., Andrade, E.R., Baruselli, P.S., Seneda, M.M. 2014. Dynamics of follicular growth and progesterone concentrations in cyclic and anestrous suckling Nelore cows (Bos indicus) treated with progesterone, equine chorionic gonadotropin, or temporary calf removal. Theriogenology 81:651-656.
Bayemi, P.H., Nsongka, M.V., Leinyuy, I., Webb, E.C., Nchadji, J.M., Cavestany, D., Bryant, M. 2015. Effect of pre-partum feed supplementation on post-partum ovarian activity, milk production and calf growth of small holder dairy Cattle in Cameroon. Tropical Animal Health Production 47:153-158.
Blanco, M., Villalba, D., Ripoll, G., Sauerwein, H., Casasus, I. 2009. Effects of early weaning and breed on calf performance and carcass and meat quality in autumn-born bull calves. Livestock Science 120:103-105.
Ciccioli, N.H., Wettemann, R.P., Spicer, L.J., Lents, C.A., White, F.J., Keisler, D.H. 2003. Influence of body condition at calving and postpartum nutrition on endocrine function and reproductive performance of beef cows. Journal of Animal Science 81:3107-3120.
Crowe, M.A., Goulding, D., Baguisi, A., Boland, M.P., Roche, J.F. 1993. Induced ovulation of the first postpartum dominant follicle in beef suckler cows using a GnRH analogue. Journal of Reproduction and Fertility 99:551-555.
Crowe, M.A., Padmanabhan, V., Mihm, M., Beitins, I.Z., Roche, J.F. 1998. Resumption of follicular waves in beef cows is not associated with periparturient changes in follicle-stimulating hormone heterogeneity despite major changes in steroid and luteinizing hormone concentrations. Biology of Reproduction 58:1445-1450.
Crowe, M.A. 2008. Resumption of ovarian cyclicity in post-partum beef and dairy cows. Reproduction of Domestic Animals 43(5): 20-28.
Crowe, M.A., Diskin, M.G., Williams, E.J. 2014. Parturition to resumption of ovarian cyclicity: comparative aspects of beef and dairy cows. Animal 8(1):40-53.
Diskin, M.G., Mackey, D.R., Roche, J.F., Sreenan, J.M. 2003. Effects of nutrition and metabolic status on circulating hormones and ovarian follicle development in cattle. Animal Reproduction Science 78:345-370.
Diskin, M.G., Kenny, D.A. 2014. Optimising reproductive performance of beef cows and replacement heifers. Animal 8(1):27-39.
Diskin, M.G., Kenny, D.A. 2016. Managing the reproductive performance of beef cows. Theriogenology 86:379-387.
Duffy, P., Crowe, M.A., Boland, M.P., Roche, J.F. 2000. Effect of exogenous LH pulses on the fate of the first dominant follicle in postpartum beef cows nursing calves. Journal of Reproduction and Fertility 118:9-17.
Funston, R.N. 2004. Fat supplementation and reproduction in beef females. Journal of Animal Science 82:E154- E161.
Galina, C.S., Rubio, I., Basurto, H., Orihuela, A. 2001. Consequences of different suckling systems for reproductive activity and productivity of cattle in tropical conditions. Applied Animal Behaviour Science 72:255-262.
Galindo-Gonzalez, S., Arthington, J.D., Yelich, J.V., Hansen, G.R., Lamb, G.C., De Vries, A., 2007. Effects of cow parity on voluntary hay intake and performance responses to early weaning of beef calves. Livestock Science110:148-153.
García-Winder M., Imakawa J., Day M.L., Zalesky D.D., Kittock R.J., Kinder J.E. 1984. Effect of suckling and ovariectomy on the control of luteinizing hormone secretion during the postpartum period in beef cows. Biology of Reproduction 31:771-778.
Granja, Y.T., Cerquera, G.J., Fernandez, B.O. 2012. Factores nutricionales que interfieren en el desempeño reproductivo de la hembra bovina. Revista Colombiana de Ciencia Animal 4(2):458-472.
Griffith, M.K., Williams, G.L. 1996. Roles of maternal vision and olfaction in suckling-mediated inhibition of luteinizing hormone secretion. Expression of maternal selectivity, and lactational performance of beef cows. Biology of Reproduction 54: 761-768.
Hernández-Martínez, J., Rebollar-Rebollar, S., González-Razo, F.J., Guzmán-Soria, E., Albarrán-Portillo, B., García-Martínez, A. 2011. La cadena productiva de ganado bovino en el sur del estado de México. Revista Mexicana de Agronegocios 15(29):672-680.
Hess, B.W., Lake, S.L., Scholljegerdes, E.J., Weston, T.R., Nayigihugu, V., Molle, J.D.C., Moss, G.E. 2005. Nutritional controls of beef cow reproduction. Journal of Animal Science 83:E90-E106.
Instituto Nacional de Estadística y Geografía (INEGI) 2018. Disponible en: http://www.beta.inegi.org.mx/app/ buscador/default.html?q=macuspana
Lents, C.A., White, F.J., Ciccioli, N.H., Wettemann, R.P., Spicer, L.J., Lalman, D.L. 2008. Effects of body condition score at parturition andpostpartum protein supplementation on estrous behavior and size ofthe dominant follicle in beef cows. Journal of Animal Science 86:2549-2556.
Lopes-Silva Filho, M., Rocha Bezerra, L., Ferreira-Silva, J. C., Póvoas Paulo Souto, F.M., Paula Oliveira, N.R., Fernandes de
Lima, P., Coutinho Bartholomew, C., Lemos de Oliveira, M.A. 2015. Influence of biostimulation and temporary weaning on follicular dynamics and pregnancy rates in Nelore cows (Bos Taurus indicus). Tropical Animal Health Production 47:1285-1291.
Lucy, M.C. 2003. Mechanisms linking nutrition and reproduction in postpartum cows. Reproduction Supplies 61:415-427.
Mackey, D.R., Sreenan, J.M., Roche, J.F., Diskin, M.G. 2000. The effect of progesterone alone or in combination with estradiol on follicular dynamics, gonadotropin profiles, and estrus in beef cows following calf isolation and restricted suckling. Journal of Animal Science 78:1917-1929.
Martins, P.G.M.A., Arthington, J.D., Cooke, R.F., Lamb, C.G., Araujo, D.B., Torres, C.A.A., Guimaraes, J.D., Mancio, A.B. 2012. Evaluation of beef cow and calf separation systems to improve reproductive performance of first-calf cows. Livestock Science 150:74-79.
Mejía-Bautista, G.T., Magaña, J.G., Segura-Correa, J.C., Delgado, R., Estrada-León, R.J. 2010. Comportamiento reproductivo y productivo de vacas Bos indicus, Bos taurus y sus cruces en un sistema de producción vaca: cría en Yucatán, México. Tropical and Subtropical Agroecosystems 12:289-301.
Mondragón, V., Galina, C.S., Rubio, I., Corro, M., Salmerón, F. 2016. Effect of restricted suckling on the onset of follicular dynamics and body condition score in Brahman cattle raised under tropical conditions Animal. Reproduction Science 167:89-95.
Montiel, F., Ahuja, C. 2005. Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: a review. Animal Reproduction Science 85:1-26.
Murphy, M.G., Boland, M.P. Roche, J.F. 1990. Pattern of follicular growth and resumption of ovarian activity in post-partum beef suckler cows. Journal of Reproduction and Fertility 90:523-533.
Pérez-Hernández, P., Sánchez-del Real, C., Gallegos-Sánchez, J. 2001. Anestro postparto y alternativas de manejo del amamantamiento en vacas de doble propósito en trópico. Investigación agraria. Producción y sanidad animales 16(2):1-14.
Pullan, N.B. 1978. Condition scoring of Fulani cattle. Tropical Animal Health Production 10:118-120.
Quintans, G., Viñoles, C., Sinclair, K.D. 2004. Follicular growth and ovulation in postpartum beef cows following calf removal and GnRH treatment. Animal Reproduction Science 80:5-14.
Quintans, G., Vázquez, A.I., Weigel, K.A. 2009. Effect of suckling restriction with nose plates and premature weaning on postpartum anestrous interval in primiparous cows under range conditions. Animal Reproduction Science 116:10-18.
Quintans, G., Banchero, G., Carriquiry, M., Lopez-Mazz, C., Baldi, F. 2010. Effect of body condition and suckling restriction with and without presence of the calf on cow and calf performance. Animal Production Science 50:931-938.
Rasby, R.J., Wettemann, R.P., Geisert, R.D., Wagner, J.J., Lusby, K.S. 1991. Influence of nutrition and body condition on pituitary, ovarian, and thyroid function of non-lactating beef cows. Journal of Animal Science 69:2073-2080.
Richards, M.W., Wettemann, R.P., Spicer, L.J., Morgan, G.L. 1991. Nutritional anestrus in beef cows: effects of body condition and ovariectomy on serum luteinizing hormone and insulin-like growth factor-I. Biology of Reproduction 44:961-966.
Salas, G., Gutiérrez, E., Ortega, R., Hernández, J. 2003. Acidos grasos no esterificados y condición corporal posparto de vacas Holstein en sistemas de producción a pequeña escala. Revista Cubana de Ciencia Agrícola 37(2):139-143.
Savio, J.D., Boland, M.P., Roche, J.F. 1990. Development of dominant follicles and length of ovarian cycles in post-partum dairy cows. Journal of Reproduction and Fertility 88:581-591.
Silveira, P.A., Spoon, R.A., Ryan, D.P., Williams, G.L. 1993. Evidence for maternal behavior as a requisite link in suckling-mediated anovulation in cows. Biology of Reproduction 49:1338-1346.
Sinclair, K.D., Revilla, R., Roche, J.F., Quintans, G., Sanz, A., Mackey, D.R., Diskin, M.G. 2002. Ovulation of the first dominant follicle arising after day 21 postpartum in suckling beef cows. Journal of Animal Science 75:115-126.
Schultz, C.L., Ely, D.G., Aaron, D.K., Burden, B.T., Wyles, J. 2005. Comparison of an early and normal weaning management system on cow and calf performance while grazing endophyte-infected tall fescue pastures. Journal of Animal Science 83:478-485.
Stagg, K., Spicer, L.J., Sreenan, J.M., Roche, J.F., Diskin, M.G. 1998. Effect of calf isolation on follicular wave dynamics, gonadotrophin, and metabolic hormone changes, and interval to first ovulation in beef cows fed either of two energy levels postpartum. Biology of Reproduction 59:777-783.
Stevenson, J.S., Jaeger, J.R., Rettmer, I., Smith, M.W., Corah, L.R. 1997. Luteinizing hormone release and reproductive traits in anestrous, estrus-cycling, and ovariectomized cattle after tyrosine supplementation. Journal of Animal Science 75:2754-2761.
Stevenson, J.S., Hill, S.L., Bridges, G.A., Larson, J.E., Lamb, G.C. 2015. Progesterone status, parity, body condition, and days postpartum before estrus or ovulation synchronization in suckled beef cattle influence artificial insemination pregnancy outcomes. Journal of Animal Science 93:2111-2123.
Vendramini, J.M.B., Sollenberger, L.E., Dubeux, J.C.B., Interrante, S.M., Stewart, R.L., Arthington, J.D. 2006. Concentrate supplementation effects on forage characteristics and performance of early weaned calves grazing rye-ryegrass pastures. Crop Science 46:1595-1600.
Vittone, J.S., Aller, J.F., Otero, G., Scena, C., Alberio, R.H., Cano, A. 2011. Destete precoz y desempeño reproductivo en vacas tratadas con progesterona intravaginal. Archivos de Zootecnia 60(232):1065-1076.
Watanabe, U., Takagi, M., Yamato, O., Otoi, T., Tshering, Ch., Okamoto, K. 2013. Metabolic profile of Japanese black breeding cattle herds: usefulness in selection for nutrient supplementation to enhance reproductive performance and regional differences. Journal of Veterinary Medical Science 75:481-487.
Waterman, R.C., Geary, T.W., Paterson, J.A., Lipsey, R.J. 2012. Early weaning in Northern Great Plains beef cattle production systems: I. Performance and reproductive response in range beef cows. Livestock Science 148:26-35.
Wettemann, R.P., Lents, C.A., Ciccioli, N.H., White, F.J., Rubio, I. 2003. Nutritional- and suckling-mediated anovulation in beef cows. Journal of Animal Science. 81(2):E48-E59.
Williams, G.L., Gazal, O.S., Guzmán-Vega, G.A., Stanko, R.L. 1996. Mechanism regulating suckling-mediated anovulation in the cow. Animal Reproduction Science 42:289-297.
Whisnant, C.S., Kiser, T.E., Thompson, F.N., Hall, J.B. 1985. Effect of nutrition on the LH response to calf removal and GnRH. Theriogenology 24:565-573.
Zieba, D.A., Amstalden, M., Williams, G.L. 2005. Regulatory roles of leptin in reproduction and metabolism: A comparative review. Domestic Animal Endocrinology 29:166-185.






.jpg&w=3840&q=75)