Introducción. Es evidente el aumento de preocupación por las prácticas ganaderas y la concienciación sobre la influencia de una dieta saludable en la salud de las personas. Por esta demanda de la sociedad, la producción animal está pasando de ser productiva ocultando problemas indeseables con antibióticos promotores de crecimiento, que pueden ser perjudiciales para la salud humana, a ser productiva a base de prevenciones y mantenimiento de la salud del animal; Este tipo de producción puede ser tan eficiente como la anterior, siempre que se complementen las dietas con productos que ayuden al animal a vencer los retos que se le presenten. Los productos más usados son probióticos, prebióticos, promotores fisiológicos, etc.
- Promotores fisiológicos. Promotores fisiológicos son los ácidos orgánicos, sus sales y las combinaciones de los mismos, que tengan un efecto positivo en la fisiología digestiva del animal por diversas razones (desarrollo del epitelio intestinal, mejora digestibilidad, aumento velocidad respuesta inmune, etc). Los compuestos que se incluyen en este tipo de aditivos se dan normalmente en el metabolismo celular, con lo que ayudan a controlar y dirigir el estado del tracto digestivo del animal. El uso de sales mejora el manejo de los productos sin afectar a la efectividad de los mismos. Los ácidos orgánicos y sus sales se han utilizado desde hace más de 30 años para reducir el contenido bacteriano y el crecimiento de hongos en materias primas y piensos compuestos, de hecho, la legislación contempla a muchos de estos como conservantes, pero los efectos positivos en la producción animal están muy bien documentados (Mallo et al. 2010).
- Nutrigenómica. La nutrigenómica está muy vinculada a la idea de adecuar la nutrición a las necesidades, no solo de las diferentes especies animales, sino incluso de los propios individuos. Es un concepto emergente, según el cual, la dieta de una persona o animal se ha de establecer a su medida, sobre la base de su propia información molecular, con la finalidad de optimizar su salud y prevenir la aparición de enfermedades. En nutrición animal es de sobra conocido que no todos los animales reaccionan de la misma manera ante un determinado alimento debido en gran parte a su particular dotación genética (Romero, 2007). Aunque un grupo de animales pertenezca a la misma raza, e incluso a la misma especie, siempre existirán pequeñas diferencias individuales en su acervo genético lo que hace que las condiciones ambientales o la composición del alimento no influyan de forma idéntica sobre la producción de dichos animales. Con la Nutrigenómica se busca establecer un conocimiento más exhaustivo del genoma para poder concebir una alimentación por grupos genéticamente homogéneos o bien una alimentación por individuo.
En los últimos años, se ha producido un importante desarrollo de ciencias como la Genómica (referida al ADN), la Transcriptómica (referida al ARN), la Proteómica (proteínas) y la Metabolómica (metabolitos), las cuales son integradas de forma conjunta por la Biología de Sistemas a través de la Bioinformática.
Aunque especialmente centrada en la nutrición humana, la Nutrigenómica es una nueva ciencia que nace como resultado de la aplicación de los métodos de trabajo de la Genómica a la Nutrición. En concreto, aúna los avances de la Genómica (identificación y efectos de los genes), la Proteómica (efectos de los genes sobre las proteínas) y de la Metabolómica (efectos de los genes sobre los metabolitos), con los de la Nutrición convencional. Sus primeros logros han sido una mayor comprensión de las repercusiones de la nutrición sobre los procesos de utilización y vías metabólicas seguidas por los nutrientes y sobre el control homeostático del organismo (Caja y Medrano, 2006).
La Nutrigenómica es, por tanto, una ciencia que busca dotar de una explicación molecular al modo en que los productos químicos ingeridos por la dieta pueden alterar el estado normal de la salud, modificando lo dictaminado por la información genética.
Promotores fisiológicos y nutrigenómica. Es importante destacar que existen muchos trabajos que demuestran el efecto beneficioso de estos productos en la nutrición de animales de granja, pero existe una delgada línea entre considerarlos de aplicación en Nutrigenómica, o simplemente considerarlos aditivos con ciertos efectos beneficiosos sobre la salud del animal o sobre su capacidad para aprovechar mejor los nutrientes de un pienso.
Por ejemplo, en un estudio realizado por Overland et al. (2007), con cerdos enteros alimentados con dietas suplementadas con diferentes ácidos orgánicos (fórmico, benzoico, butírico) se concluía que el consumo de estos ácidos disminuía el número de coliformes, enterococos y bacterias ácido lácticas en el tracto gastrointestinal, y no encontraron diferencias en el nivel de escatol (compuesto responsable del olor sexual de la carne de cerdo) ni en el colon ni en el tejido adiposo. Seguramente guarda relación con la expresión de algún gen, especialmente la producción de escatol, pero no era objeto de este ensayo.
En una prueba realizada por Smulikowska et al (2009) en pollos se demostró que una mezcla de ácidos orgánicos y sales (fumárico, formiato de calcio y propionato de calcio y sodio) afectaba positivamente a la morfología del intestino delgado, aumentando la altura de vellosidades y la profundidad de las criptas. Una vez más, la relación con la Nutrigenómica no está claramente demostrada. Otro caso parecido en cabras se da en el estudio de Ishiwata (2000) donde concluyen que el propionato y el butirato en cultivo in vitro, pueden inhibir la GH (hormona de crecimiento) y la expresión del ARNm en células de la hipófisis anterior de cabras.
- Ácido propiónico. En alimentación animal, se utiliza habitualmente como conservante para el pienso porque inhibe el crecimiento de hongos y algunas bacterias.
Ácido propiónico en terneros. La suplementación de la leche con propionato de cromo y de calcio en terneros no afecta a los rendimientos productivos, sin embargo el propionato de cromo puede producir un aumento de la secreción de insulina en sangre (Bunting et al., 2000), con un índice de desaparición de glucosa de 0.013 unidades en el caso del grupo control y 0.019 en el caso de los animales suplementados con 0.5 mg/kg de propionato de cromo en el pienso.
Ácido propiónico en vacas. Bradford et al. (2006) estudiaron la inclusión de propionato en dietas de vacas lecheras y vieron que aumentaba la síntesis de leptina en la sangre (hormona que regula la ingestión de alimento). Defrain et al (2005) demostraron que la suplementación de la dieta con propionato de sodio y calcio no afecta a la producción de leche pero sí a su composición: la cantidad de grasa fue mayor y MUN (nitrógeno ureico en leche) se redujo en el caso de la dieta sin propionato (tabla 1).

- Ácido butírico. El ácido butírico es un líquido corrosivo, muy volátil a temperatura ambiente y de olor fuerte. Por su difícil manejo en las fábricas de piensos se comercializa principalmente en forma de butirato sódico. El ácido butírico juega un papel muy importante en la fisiología y metabolismo ruminal, intestinal y de las mucosas del aparato digestivo en general. Además de ser una fuente de energía ideal para los enterocitos, se ha demostrado que previene enfermedades relacionadas con el aparato digestivo.
El ácido butírico tiene efectos a nivel molecular, celular y epitelial, actúa como inhibidor de las histona deacetylasas (HDACs); influye en la expresión de ciertos genes a nivel celular; afecta al crecimiento, diferenciación y apoptosis celular en el epitelio intestinal; reduce la inflamación epitelial (disminuyendo la expresión de citoquinas como IL-6 e IL-1β). Ademαs, el butirato estimula la secreciσn de enzimas digestivas en el tracto gastrointestinal y puede aumentar el consumo por la activaciσn de receptores especνficos (Galfi, 2010).
En la producción animal, diferentes estudios han demostrado que la utilización de butirato como aditivo para piensos es muy útil ya que permite el control de patógenos, aumenta la digestibilidad de la ración, la tasa de crecimiento, promueve la producción de citoquinas y disminuye el estrés oxidativo, incluso en dosis muy bajas. Los efectos son en su mayoría notables en neonatos y antes del destete de los lechones y terneros (Guilloteau et al., 2010 a), aunque también se ha demostrado su efectividad en cuanto a parámetros zootécnicos en animales adultos y en aves.
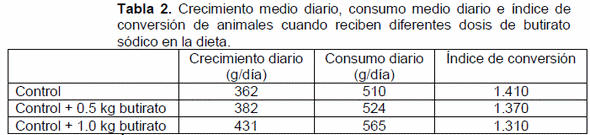
Ácido butírico en porcino. Numerosos estudios han demostrado que su inclusión en la dieta produce mejoras significativas en varios aspectos. Mejora el crecimiento y la morfología intestinal: aumenta la altura de vellosidades y profundidad de las criptas en el intestino delgado, produciendo un incremento de la superficie de absorción de nutrientes y aumentando su digestibilidad. Además, interviene en el sistema inmunológico influyendo en la proliferación de células mononucleares de la sangre (linfocitos y monocitos), componentes clave en el sistema inmune (Thomas et al., 2006). También disminuye el factor de necrosis tumoral alfa y los niveles de interleucina 6 (factores relacionados con la población de células del sistema inmunitario), aumenta la producción de citoquinas (proteínas que regulan la función de otras células) y reduce la concentración de bacterias intestinales como E.coli y Clostridium (Lu et al., 2008). Kotunia et al. (2004) demostraron también que el butirato sódico en dietas de lechones neonatos aumentaba la concentración en plasma de polipéptidos y colecistoquinina, hormona que participa en la regulación de la digestión. También Mazzoni et al. (2008), diferenciando entre su utilización en la alimentación pre y post destete, observaron un aumento del número de células parietales, independientemente del periodo de administración, pero después del destete vieron además un aumento del número de células entero-endocrinas, así como de la somatostatina (hormona producida por las células del páncreas que interviene en la regulación de la glucemia) y tendió a aumentar el ARNm de la gastrina. Todos estos efectos contribuyen a que los animales aprovechen mejor los nutrientes del pienso y sean más resistentes a enfermedades. Como ejemplo, en esta tabla se dan los resultados productivos de lechones de 6.7 kg de peso inicial durante los primeros 30 días post-destete, cuando se le suministraba en la ración butirato sódico a 2 dosis diferentes (Lu et al. 2008) (tabla 2).
Ácido butírico en avicultura. La investigación con butirato sódico en pollos se centra principalmente en su utilización para la prevención de infecciones. En un estudio con pollos a los que se les inoculó E.coli se produjo una reducción del nivel de interleucina y del factor alfa de necrosis tumoral en los pollos alimentados con butirato sódico, igual que en el caso de lechones, además de un aumento de actividad de la catalasa y la peroxido dismutasa (enzima antioxidante). En los animales infectados no se produjo disminución de la ganancia de peso ni del consumo, por lo que se concluye que el butirato podría mejorar el rendimiento en pollos sometidos a estrés (Zhong et al., 2011).
Es muy interesante observar como el uso de promotores fisológicos adecua la transcripción genética de los animales a la situación en la que se encuentran. Así por ejemplo, en el caso de que los animales se estén enfrentando a un desafío infeccioso, la respuesta inmune se potencia cuando los animales reciben butirato sódico protegido (Mallo et al. 2011), el nivel de Interleuquina 2, un inmunoestimulante, aumenta, y el factor de tumoración necrótica inducida por polisacáridos alfa (LITAF) disminuye; mientras que estos cambios no se observan cuando los animales no se enfrentan a un desafío. Esta adaptación se ve potenciada si se combina el butirato sódico con aceites esenciales en el promotor fisiológico; en ese caso, disminuye además la transcripción del factor de crecimiento vascular endotelial TNFSF15, relacionado con la apoptosis de las células defensivas y con la inflamación epitelial.
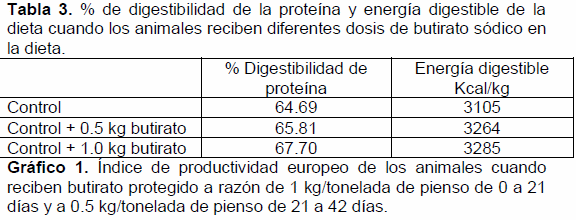
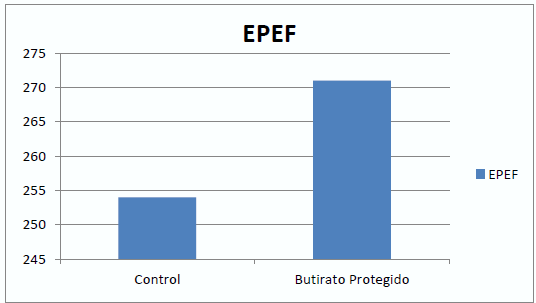
De la misma manera que en porcino, el butirato afecta al desarrollo de las vellosidades intestinales (Mallo et al., 2010), y este mayor desarrollo afecta a la digestibilidad de la dieta (Mallo et al., 2011) (tabla 3). Y al crecimiento de los animales (Mallo et al., 2010) (gráfico 1).
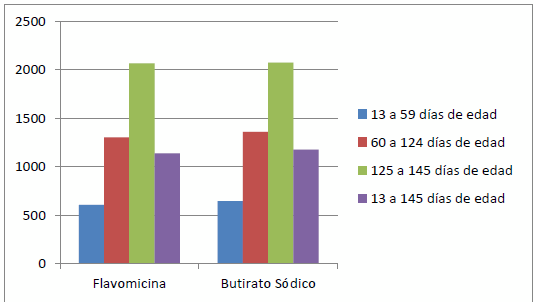
Ácido butírico en terneros. En rumiantes, las investigaciones van dirigidas a cambios ocurridos en el rumen, páncreas y secreciones plasmáticas de hormonas o enzimas. Así, Gorka et al. (2009) demostraron que la inclusión de butirato de sodio en la leche en terneros neonatos no afectaba a la tasa de crecimiento pero sí que reducía la pérdida de peso respecto a la dieta control. También aumentaba el peso del retículo-rumen y la longitud y anchura de sus papilas. Además, observaron una mayor cantidad de GLP-2 en sangre (hormona que se encarga de la secreción de insulina por el páncreas y la disminución en los niveles de glucosa en sangre). Otros estudios recientes demuestran que la ingesta de butirato de sodio estimula las secreciones del páncreas, en particular la quimiotripsina (enzima proteolítica), la lipasa (Guilloteau et al., 2010 b) y la elastasa (Guilloteau et al., 2009). Esto puede explicar el aumento de la digestibilidad de algunos nutrientes al incluir butirato en la alimentación. Además, aumentó ligeramente la concentración plasmática de secretina (hormona gastrointestinal) y coleocistoquinina. También se incrementó el nivel de HSP (proteínas de choque térmico que producen las células en situación de estrés) en el estómago y el colon, lo que podría significar un estímulo de la citoprotección frente a úlceras gástricas que se producen con frecuencia en terneros; De esta manera se observa como animales que reciben butirato sódico en la dieta se desarrollan mejor que animales que reciben flavomicina (Guilloteau et al., 2009) (gráfico 2).
Gráfico 2. Crecimiento de animales recién destetados cuando reciben butirato sódico en la ración o flavomicina.
Ácido butírico en vacas. Existen varios estudios en los que se ha evaluado el papel del butirato sódico en la infección por Staphylococcus aureus, (responsable de la mastitis en bovinos) en el epitelio mamario de vacas y en la expresión de genes de péptidos antimicrobianos. En concreto, Ochoa-Zarzosa et al. (2009) concluyeron que el butirato puede reducir la internalización bacteriana y es capaz de regular la expresión de péptidos antimicrobianos traqueales, b-defensinas (proteína que actúa como antibiótico natural), inducción del ARNm de la óxido nítrico sintasa, así como la producción de óxido nítrico. Además, observaron una acetilación de la histona H3 en las células epiteliales mamarias, lo que indica que el butirato de sodio podría ser eficaz para modular la expresión génica inmune innata de la glándula mamaria y mejorar la defensa contra la infección bacteriana.
Los efectos del butirato sódico en la expresión génica fue estudiado también por Li et al (2006), en concreto en las células epiteliales del riñón de bovinos. El butirato afectó a 450 genes, la mayoría de ellos asociados con el control del ciclo y la apoptosis celular. Un estudio previo (Li and Elsasser., 2005) también demostró este hecho, mostrando el efecto pleiotrópico del butirato.
Conclusión. El uso de promotores fisiológicos en la nutrición de animales, tanto monogástricos como rumiantes, para modificar la expresión de determinados genes, y con ello los niveles de enzimas y hormonas principalmente, puede resultar una herramienta muy útil para conseguir una mejor adaptación de determinados animales a sus respectivas dietas, mejorando su sistema inmunitario y digestivo al mismo tiempo. De esta manera los animales se adaptan mejor a las dietas y confrontan los desafíos inmunológicos con un mejor estado general, aumentando la posibilidad de que expresen su potencial genético y mejorando la productividad.
Bibliografía. Bradford BJ et al. 2006. Domestic Animal Endocrinology 30: 65-75; Buntig LD et al. 2000. J. Dairy Sci. 83: 2491-2498; Defrain JM et al. 2005. J. Dairy Sci. 88: 983-993; Galfi P 2010. I Jornada Técnica Avicultura 1-12; Gorka P 2009. Journal of Physiology and Pharmacology 60/3: 47-53; Guilloteau P 2010. Nutrition Research Reviews 23:366-384; Guilloteau P 2010. J. Dairy Sci. 93: 5842-5850; Guilloteau P 2009. J. Dairy Sci. 92: 1038-1049; Li CJ and Elsasser TH 2005. Journal of Animal Science 83: 89-97; Li RW et al. 2006. BMC Genomics 7:234; Lu JJ et al. 2008. Journal of Animal and Feed Sciences 17: 568-578; Mallo JJ et al. 2011. International Poultry Scientific Forum 2011; Mallo JJ et al. 2010. XIIIth European Poultry Conference; Mallo JJ et al. 2011. Congreso AECA 2011









