Mediciones de eficiencia nutricional en Pollos Parrilleros de acuerdo al objetivo de producción
Publicado: 8 de marzo de 2006
Por: Julian Melo, MV, Ms. Sc.Dpto. Producción Animal, Facultad de Ciencias Veterinarias, Univ. de Buenos Aires
Desde un punto de vista energético, la acumulación de tejidos como causante del aumento de peso es consecuencia del exceso de energía consumida por sobre los requerimientos de mantenimiento (Soller & Eitan, 1984). Pudiéndose expresar también, a nivel celular, como el momento en que la síntesis de macromoléculas supera a la degradación de las mismas.
La tasa de retención de proteínas y grasas, como asi también su distribución, depende de variables propias del animal (peso, ganancia de peso, edad, sexo, biotipo, estirpe) como de variables externas (alimentación, clima, manejo, estado sanitario). De acuerdo a Webster (1989), si las condiciones externas no son limitantes, el organismo tiene como objetivo acumular en el tiempo una cantidad determinada de proteína tisular, que determina el tope biológico de crecimiento. Este límite se considera determinado genéticamente. El mismo autor (Webster, 1989) también señala que las variables que afectan la respuesta animal lo hacen en mayor magnitud sobre la cantidad de grasa que sobre la de tejido magro.
La composición química corporal, excluyendo las plumas, cambia a medida que el animal crece. Los cambios mas importantes son el aumento del contenido de grasa y la disminución de agua. La composición de la fracción seca libre de grasas se mantiene constante con el crecimiento del ave (Gous, 1998). En promedio la composición química de un pollo comercial (incluyendo plumas) a los 40 días es: 65 % agua, 19 % proteína, 13 % lípidos y 3 % cenizas.
La mayoría de la materia seca de los músculos, que constituye tan solo un 25% del peso húmedo, se encuentra en forma de proteína (Soller y Eitan, 1984). Este tejido contribuye con el 51 % de la proteína total del ave, mientras que el tejido conectivo lo hace con el 23 %, el hígado con el 5%, el plasma con el 3% y el tracto gastrointestinal con el 5% (Klasing, 1993). La síntesis y degradación proteica son procesos energéticamente caros, siendo baja la eficiencia de su deposición. Es decir, que una gran cantidad de proteína se está depositando mientras otro tanto se esta degradando. Pollos seleccionados por una conversión eficiente de la energía dietaria en proteína corporal lograron altas tasas de deposición proteica con bajas tasas de degradación, en comparación con una población control (Klasing et al., 1987).
El tejido adiposo tiene una cantidad de grasa, agua y una distribución que depende de la dieta, del peso, de la edad y de la raza, biotipo o estirpe. Su composición varía entre un 70 a un 75 % de grasa, 20 a 25 % de agua y 6 a 7 % de proteína, estando distribuida principalmente en tres regiones: piel y subcutáneo (45 %), panículo abdominal (23 %) y entre las fibras musculares (31 %), de acuerdo a las determinaciones realizadas por Carden et al.(1981). El coeficiente de variación para grasa abdominal se encuentra entre un 25 a un 30 %, en comparación con el 15 a 20 % para grasa total, y en total contraposición con los coeficientes de variación para agua, proteína y cenizas (aproximadamente un 2, 3 y 8 %, respectivamente) (Pym, 1988).
Eficiencia Nutricional:
Metabolismo Energético: La energía que se necesita para la síntesis de tejidos y para cualquier otro proceso metabólico que precise energía, es el resultado de la oxidación continua de substratos orgánicos que aportan tanto el alimento, por medio de sus nutrientes, como los tejidos, que se degradan continuamente. Por lo tanto, si la alimentación es insuficiente para mantener al animal, aumenta la degradación tisular y produce una pérdida de peso en el mismo (Di Marco, 1993). La demanda de energía depende del gasto que se realice en los distintos tejidos, lo cual está influenciado por las funciones metabólicas que lleven a cabo los mismos (Webster, 1989). Las funciones metabólicas que demandan energía se agrupan en tres categorías: trabajo fisiológico, transporte de iones de Na+/K+ y biosíntesis de proteínas y grasas. Refiriéndose la primera, por ejemplo, a la actividad del corazón, pulmones y peristaltismo intestinal, y la segunda, a la actividad de las bombas de Na+ y de K+ para mantener el potencial de membrana. En general, dentro del total de energía demandada, la mayor proporción demandada la da el transporte de iones, luego la síntesis protéica, y por último el trabajo fisiológico. En cada tejido predomina alguna de estas funciones, y ello determina las distintas demandas energéticas de los tejidos. Aquellos que cumplen funciones de servicio para el resto del organismo ( cardíaco, pulmonar, hepático, gastrointestinal, renal ) se los ha denominado "supletorios", mientras que los que principalmente estan orientados a una funcion de biosintesis ( muscular, graso, plumas), se los ha denominado "demandantes" (Susbilla et al., 1994). Los primeros, que representan un bajo porcentaje del peso vivo, tienen una alta proporción del flujo sanguíneo y una alta intensidad metabólica, es decir, una importante tasa de síntesis y degradación con un importante gasto en el transporte de iones; siendo el gasto energético por unidad de peso 45 veces mayor en el riñón que en el tejido muscular (Di Marco, 1993). La mayor proporción de peso de órganos digestivos, en parte explica la mayor demanda energética por unidad de peso de los animales jóvenes y de las estirpes genéticas no seleccionadas por eficiencia nutricional y ganancia de peso (Melo et al., 1999; Miquel et al., 1998). Es decir, que a mayor peso disminuye el gasto energético por unidad de peso, lo cual esta contemplado por el concepto de peso metabólico ( Kg0.75 ).
Partición de la energía: Clasicamente, a la Energía Metabolizable (EM), se la puede descomponer en Energía de Mantenimiento (MA), Incremento calórico de Mantenimiento (ICm), Energía Retenida (ER) e Incremento Calórico de Producción (ICp). Se acepta que solo se pueden incluir los términos ER e ICp cuando hay excedente de EM, que entonces se puede destinar a la producción (síntesis de proteína y grasa). También se puede particionar a la energía desde el punto de vista de las funciones metabólicas, ya que es metodológicamente imposible poder discernir entre los procesos de mantenimiento y producción. Los modelos que integran aspectos metabólicos con la respuesta animal utilizan esta partición y calculan el costo de mantenimiento en el proceso iterativo como el punto en que el balance energético es nulo (Di Marco, 1993). La definición clásica de mantenimiento dice que el animal esta en ese estado cuando no gana ni pierde peso, lo cual, desde el punto de vista fisiológico no estaría reflejando casos en los cuales el tejido adiposo es reemplazado por agua, por ejemplo. El mantenimiento es un costo energético variable afectado por la composición corporal y por la proporción de tejidos metabólicamente activos del animal.
Formas de expresar la eficiencia:
Eficiencia energética de la síntesis: Es la energía utilizada en la síntesis de tejidos en relación a la energía total demandada por el organismo. Siendo la formación de tejidos, en general, una pequeña fracción de la ya pequeña que se utiliza en la síntesis de macromoléculas (15-20 %), de acuerdo a Webster (1989). Los tejidos de los órganos internos y vísceras son los principales responsables del gasto de energía a nivel tisular, mientras que a nivel celular lo es el transporte de iones. De acuerdo al mismo autor estas son las principales causas de diferencias en eficiencia entre individuos o biotipos. En términos fisiológicos, se sintetiza tejido mas eficientemente cuanto menor es el peso de los tejidos de alta intensidad metabólica y cuanto menor es el gasto en el transporte de iones, ya que de esa manera hay un menor gasto de mantenimiento (Di Marco, 1993).
Eficiencia en la retención de tejidos: Se asume que las ganancias de peso, tanto del tejido magro como del graso, están directamente relacionadas con un balance energético positivo. Por ello, para que haya crecimiento tiene que haber un exceso de consumo de energía, lo cual determina que el consumo sea uno de los principales causales de la retención energética. Sin embargo, no tiene el mismo costo energético la deposición de un gramo de tejido graso que la de un gramo de tejido magro, ya que si bien demanda la misma energía depositar un gramo de proteína que de grasa, el tejido graso tiene un 20 % de agua y el magro un 80 %, lo cual lleva a que se depositen 0,4 gr y 0,1 gr de tejido magro y graso, respectivamente, por cada kcal de EM (Webster, 1980). Esto determina que a mayor deposición de tejido magro mas eficiente será el individuo.
Eficiencia en la utilización de la EM: Depende del destino de la energía, mantenimiento o producción, y del tipo de tejido que se produzca. El calculo de esta eficiencia asume que el metabolismo de ayuno y el costo de mantenimiento permanecen constantes en cualquier nivel de consumo. No existe duda de que algunos genotipos utilizan la EM mas eficientemente que otros, pero todo indicaría que las diferencias genéticas serían determinadas no por diferente utilización de la energía de producción sino por la de mantenimiento. Estas diferencias se podrían dar por alguno o varios de los factores que se involucran en el mantenimiento del ave, siendo cuatro de los mismos de particular interés: la actividad física, el mantenimiento de la temperatura corporal, el mantenimiento de los tejidos corporales y la tasa metabólica basal (Gous, 1999).
Eficiencia en la conversión de alimento (kg de alimento consumido / kg de peso ganado): Es la mas importante desde el punto de vista productivo. A mayor consumo mayor eficiencia de conversión, ya que se diluye el costo de mantenimiento, por lo cual, los principales determinantes de la eficiencia de conversión van a ser el nivel de consumo, el costo de mantenimiento y el tipo de tejido retenido.
Consumo residual: Debido a que el alimento consumido es utilizado tanto para mantenimiento como para ganancia de peso, se podría afirmar que la Tasa de Conversión es una medida 'bruta' y no 'neta' de la eficiencia. Por otro lado, debido a que el incremento del peso corporal por unidad de alimento consumido es una función decreciente del peso vivo, la ganancia de peso debería corregir las mediciones de consumo a una misma edad (Chambers y Lin, 1988). Los modelos de regresión que incluyen el peso vivo como covariable son usualmente utilizados para ajustar el consumo por diferencias en peso vivo. Esta metodología realiza el ajuste removiendo, al menos, el efecto de las diferencias en el peso. Chambers y Lin (1988) hallaron que la variación remanente luego de corregir el consumo por peso vivo y por ganancia de peso estuvo entre 2 y 9 %, demostrando que las diferencias en consumo entre individuos se debían en gran medida a la tasa de crecimiento. A este remanente de cada medición de consumo con respecto a lo predicho en función de su peso y tasa de crecimiento se lo define como Consumo Residual. Las posibles razones para explicar la variación remanente serían, por un lado, las diferencias entre animales para composición corporal, mantenimiento u otro proceso que demande energía, y, por otro lado, los errores en la predicción y en la medición del consumo (Kennedy et al, 1993).
Medición de la eficiencia en la retención diferencial de tejido
La medición del metabolismo energético y nitrogenado utilizando jaulas individuales no calorimétricas ha sido descripta por varios autores. Parte de los mismos utilizaron como técnica la faena comparativa (Alleman et al., 2000; Geraert et al., 1988; 1990; 1993; Lippens et al., 2002; Melo, 2002; Melo et al., 2004), mientras que otros utilizaron como técnica la recolección de heces individuales (Pym & Farell, 1977; Sklan & Plavnik, 2002). La técnica de faena comparativa precisa la faena previa de una muestra representativa de animales para obtener una estimación de la composición corporal inicial de los animales que continuarán el ensayo. También se obtiene la medición del consumo individual y del peso vivo.
Al finalizar el período de prueba se sacrifican todos los animales que se encontraban en las jaulas. A cada pollo sacrificado se lo despluma y pasa por una picadora hasta homogeneizar el producto final. Del picado final de cada pollo se toman muestras para determinar en cada una de ellas la materia seca, el extracto etéreo y la proteína cruda, de acuerdo a métodos de análisis descriptos por la A.O.A.C. en 1980. La diferencia entre la composición química estimada para el inicio y la obtenida al final del ensayo se utiliza para estimar la grasa y proteína retenida por cada individuo.
La cantidad de energía retenida como proteína y grasa se estimó multiplicando a los gramos retenidos por 5,66 y 9,37 Kcal, respectivamente. Siendo la energía retenida total la sumatoria de las energías retenidas como proteína y como grasa. También es factible estimar la eficiencia proteica debido a que se cuenta con la medición de la cantidad de proteína consumida y retenida (Ret./Cons.).
Resultados obtenidos en distintos ensayos
Los valores medios de la fracción de energía retenida como proteína y como grasa que se obtuvieron varían de acuerdo a la dieta, edad, estirpe y temperatura que se utilizaron para la realización de los ensayos. El incremento proteico en la dieta bajo dietas isoenergéticas no siempre significó un aumento de la media de fracción de energía retenida como proteína, aunque en una línea magra bajo condiciones de termoneutralidad siempre hubo una respuesta positiva al agregado de proteína a la dieta hasta un 23 % (Geraert et al., 1990; 1993). Como era de esperar el aumento de la edad influye negativamente en la fracción de energía retenida como proteína (Geraert et al., 1988), al igual que una temperatura constante de 32 ºC (Geraert et al., 1993), mientras que temperaturas bajas (10 ºC) no dieron resultados distintos a los obtenidos bajo termoneutralidad (Geraert et al., 1988). Estirpes genéticas seleccionados por mayor o menor grasa abdominal dieron como resultado una variación consecuente (Geraert et al., 1988, 1990, 1993), mientras que Jorgensen et al. (1990) encontraron diferencias significativas entre líneas seleccionadas por conversión y por ganancia de peso, siendo esta última de menor proporción de energía depositada como proteína. En estirpes no seleccionadas, Melo et al.(2004) encontraron resultados intermedios a los de las líneas grasa y magra en condiciones de elevada temperatura (Geraert et al., 1993), lo cual era esperable ya que la primera evaluación fue llevada a cabo en verano.
Los valores medios de eficiencia proteica también se ven afectados por una elevada temperatura ambiental, mientras que la disminución a 10 ºC no llevaría a diferencias con la termoneutralidad, de acuerdo a lo informado por Cahaner y Leenstra (1992), Geraert et al. (1988, 1993) y Melo et al.(2004). Como era de esperar la edad también afecta negativamente a la eficiencia proteica (Geraert et al., 1988), mientras que las diferencias entre genotipos son las mismas que las señaladas para la fracción de energía retenida como proteína. Donde se observa una gran diferencia con respecto a la mencionada fracción es en la disminución de la eficiencia proteica con el incremento de proteina cruda en la dieta en un rango entre 13 y 25 % (Geraert et al., 1990, 1993; Sklan & Plavnik, 2002).
Parámetros productivos y otras medidas de eficiencia
En trabajos previos se ha demostrado que las diferencias en el contenido de grasa de dos estirpes genéticas seleccionadas en forma divergente por proporción de grasa abdominal no se pudieron explicar enteramente por diferencias en el consumo o pérdida de energía (Leclercq y Saadoun, 1982; Geraert et al., 1988; Geraert et al., 1990; Geraert et al.., 1993). Sin embargo, si se encontraron diferencias significativas en la fracción de energía retenida como proteína o como grasa, siendo aumentada la fracción retenida como proteína y disminuida la fracción retenida como grasa en la estirpe magra, y viceversa. Melo et al.(2004) encontraron una mayor variabilidad para la fracción proteica (ERP/ERT) que para la fracción grasa (ERG/ERT), lo cual parecería indicar una contraposición con la habitualmente mayor variabilidad de las proporciones de tejidos grasos con respecto a los magros y también en contraposición a la mayor variabilidad de los porcentajes de extracto etéreo con respecto a los porcentajes de proteína bruta corporal. Sin embargo, ambas variables se comportan complementariamente y si se busca a los individuos de mayor proporción de energía retenida como proteína van a ser los mismos individuos que tienen la menor proporción de energía retenida como grasa, lo cual no cambiaría la decisión que se tome con respecto a diferentes genotipos o dietas.
A pesar de existir escasa bibliografía referente a las correlaciones entre las variables de eficiencia en la retención diferencial de tejido y las variables de producción, si se realizaron análisis de varianza para comparar a las líneas magra y grasa mencionadas en el párrafo anterior para estos dos conjuntos de variables. Como ya habíamos señalado, estas líneas se diferenciaron en forma significativa (p<0,05) en las variables de eficiencia en la retención diferencial de tejido, lo cual nos da una noción de la magnitud en que se asociaron con algunas de las variables de producción. Melo et al. (2004) hallaron una correlación cercana a cero entre el peso vivo de pollos de 49 días de una estirpe de crecimiento lento y las fracciones de energía retenida, en concordancia con la ausencia de diferencias significativas (p>0,05) entre los pesos de las líneas magra y grasa encontrados por Geraert et al. (1988;1990) para las 7 semanas de vida. Sin embargo, los citados trabajos muestran que a mayor edad el peso vivo si parecería tener algún tipo de asociación con la fracción de energía retenida, ya que de acuerdo con las correlaciones y coeficientes de regresión obtenidos los animales de mayor fracción proteica retenida fueron de menor peso vivo a los 79 días que los animales de mayor fracción grasa retenida, en animales de crecimiento lento (Melo, et al., 2004), mientras que Geraert et al. (1993) en contraposición a estos resultados informa un mayor peso vivo de la línea magra con respecto a la grasa a los 63 días. En ese trabajo las líneas se diferencian en forma significativa por una mayor ganancia de peso para la línea magra, siendo la misma de mayor ganancia de proteína a pesar de tener un menor consumo de proteína. Esta diferencia deja claro que la mayor ganancia de peso de un individuo no significaría mayor ganancia de proteína en todos los genotipos y condiciones (Gráfico 1).
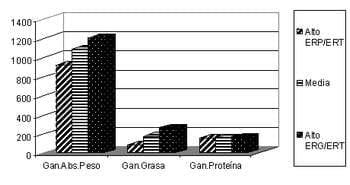
Gráfico 1: Ganancia de peso (gr) total y por componente químico para los 20 pollos con mayor energía retenida como proteína (ERP/ERT) y los 20 con mayor energía retenida como grasa (ERG/ERT) entre los 49 y los 79 días de vida (Melo et al., 2004).
En la mayoría de los casos el consumo voluntario de los animales se expresó en el mismo sentido que la ganancia de peso, siendo los individuos de menor fracción grasa retenida los que presentaron menor consumo, de acuerdo a las correlaciones y coeficientes de regresión obtenidos por Melo et al.(2004) y en conformidad con lo descrito por Geraert et al.(1988, 1990, 1993) en sus trabajos. Las diferencias en consumo parecerían ser explicadas en gran proporción por el peso vivo y por la ganancia de peso debido a que no hubo diferencias en el consumo residual de estos animales y, por ende, en el consumo corregido por el peso y la ganancia de peso (Gráfico 2). Es decir que la mayor parte del exceso de consumo de los animales de menor fracción proteica retenida se debe a las diferencias en ganancia de peso y peso vivo que a diferencias en su composición química.
El menor consumo de los animales de alta fracción proteica en una población de crecimiento lento no alcanzó para presentarlos como de mejor conversión debido a que también fueron animales de menor ganancia de peso (Melo, et al., 2004). Esto se contradice con lo hallado para las líneas grasa y magra de Geraert et al..(1988,1990,1993), que tuvieron conversiones alta y baja, respectivamente y diferentes en forma significativa (p<0,05).
Eficiencia Proteica: Considerando que las líneas magra y grasa utilizadas por Geraert et al. (1988,1990,1993) también difirieron en eficiencia proteica en forma significativa (p<0,05), se observó que las mismas siempre difirieron en conversión, como resultado de diferencias en consumo y ganancia de peso (Geraert et al., 1993) o solo de Consumo (Geraert et al..., 1990), en concordancia con el sentido de las correlaciones (r=0,47) presentadas por Melo, et al. (2004). La diferencia en eficiencia proteica entre los animales no parecería deberse principalmente a un bajo peso inicial, a pesar que ello determinó la diferencia de ganancias de peso relativa, ya que el consumo residual negativo para los animales de mayor eficiencia proteica podría estar indicando la deposición de una mayor fracción proteica de tejido, debido a que es la mas eficiente de realizar (Soller y Eitan, 1984). Lo cual también se sustenta en que el porcentaje de variación de la eficiencia proteica explicado por el peso vivo a los 49 días que informan Melo, et al., (2004) fue de solo un 9 %, mientras que la proteína bruta corporal lo hizo en un 42 %, de acuerdo al R2 de los modelos de regresión simple respectivos. La diferencia en composición química corporal era preexistente a la medición de la eficiencia proteica y se mantuvo hasta los 79 días, independientemente del peso vivo, ya que a los 49 días había un 45 % de gramos de proteína con respecto al total de gramos retenidos para los 20 animales de mayor eficiencia proteica y 40 % para los 20 pollos de menor eficiencia proteica, diferencia que se mantuvo a favor del primer grupo a los 79 días (Melo, et al.,2004).
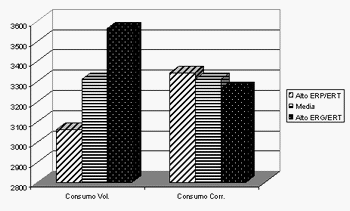
Gráfico 2: Consumo (gr) Voluntario y Corregido por peso y ganancia de peso para los 20 pollos con mayor energía retenida como proteína (ERP/ERT) y los 20 con mayor energía retenida como grasa (ERG/ERT) entre los 49 y los 79 días de vida (Melo et al., 2004).
El peso vivo inicial mas bajo para los animales de alta eficiencia proteica podría observarse por la tendencia de los genotipos mas magros a tener una curva de crecimiento mas lenta al inicio, de acuerdo a lo explicitado por Geraert et al.. (1993), en el sentido que los animales magros mostraron menor crecimiento inicial que los grasos pero a los 63 días de vida obtuvieron mayor ganancia de peso, mejor conversión y mayor peso vivo con un menor consumo voluntario (p<0,05). Esto podría estar sugiriendo que los animales de mayor eficiencia proteica serían animales de mayor peso a la madurez, lo cual significaría un mayor requerimiento en los reproductores, pero que al conjunto del sistema le sería mas que compensado por un aumento de eficiencia en el crecimiento de una proporción muy grande de animales en relación a la cantidad de reproductores. Las líneas seleccionadas por Jorgensen et al... (1990) y por Leenstra y Pit (1987) para la mejora de la conversión obtuvieron una mejora correlacionada de la eficiencia proteica (Cahaner y Leenstra,1992), en concordancia con las correlaciones y el signo de los coeficientes de regresión que obtuvieron Melo, et al. (2004).
Discusión
Pym (1996) obtuvo el mismo resultado en conversión para dos estirpes que diferían en un 33 % en grasa, y que habían sido seleccionadas por menor grasa una y por mayor ganancia de peso la otra. En ese sentido, parecería que las variables de producción no tendrían claros efectos sobre las variables de eficiencia en la retención diferencial de tejido. La existencia de correlación genética entre la proporción grasa y la conversión, que no es alta pero es negativa, explica la obtención de respuestas correlacionadas (Jorgensen et al..., 1990; Pym y Solvyns, 1979; Geraert et al.,1990,1993). Sin embargo, existirían otras posibilidades para mejorar la eficiencia nutricional en las cuales no estaría comprendido el cambio en el tipo de tejido retenido, como sería el aumento de la cantidad de energía metabolizable que se puede obtener de la dieta (Jorgensen et al. 1990) o el aumento del turn-over proteico, o la disminución del requerimiento de energía para el mantenimiento, lo cual parecería sugerir que sería necesaria alguna medición de composición corporal directa si se quiere mejorar la eficiencia nutricional por medio de la eficiencia en la retención diferencial de tejido, si fuera el caso donde la composición corporal es parte del beneficio económico. Los bajos coeficientes de determinación obtenidos para la predicción de alguno de los parámetros de eficiencia en la retención diferencial de tejido por medio de variables de producción, aunque se utilizaran conjuntamente (Melo, et al., 2004), parecería indicar una mediana o baja asociación entre los mismos, lo cual sustentaría lo arriba señalado. Esto, sin embargo, no impediría hacer algunas observaciones sobre como se podrían utilizar las variables de producción si se pretende mejorar la eficiencia nutricional por medio de la mejora en la eficiencia en la retención diferencial de tejido. De los resultados obtenidos parecería que existen variables de eficiencia en la retención diferencial de tejido que en forma directa no darían bajos pesos finales, ya que los animales de alta eficiencia proteica son de bajo peso inicial y alcanzan el peso promedio al llegar la edad de faena, obteniendo una ganancia de peso por encima de la media (Grafico 3).
Esto plantearía un crecimiento lento al inicio y aumentar rápidamente en la última etapa previa a la faena, como fuera ya sugerido por varios autores (Jones & Farell, 1992; Leeson & Zubair, 1997; Lippens et al., 2002). Este tipo de curva de crecimiento permitiría un menor gasto de mantenimiento en los animales y una menor deposición de grasa relativa al total de energía retenida, sin dejar de resignar el aumento en la variable que mayor incidencia tiene en el beneficio económico del productor, como es el peso vivo final.
En todos los casos los animales de mayor eficiencia del trabajo realizado por Melo, et al. (2004) tuvieron un consumo voluntario inferior a la media (Grafico 4). Para agregar sustento a lo escrito hasta aquí, Webster (1989), en cierta medida, también recomienda que se considere a mediciones como la eficiencia proteica o la fracción ERP/ERT para la determinación de individuos mas eficientes en la retención de tejidos pero que no resignen peso final, ya que considera que la mejor forma de hacerlo es considerar a la energía retenida como proteína en función del consumo como criterio para lograrlo, lo cual permitiría elegir a los animales de mayor ganancia de tejido magro con el menor consumo posible.
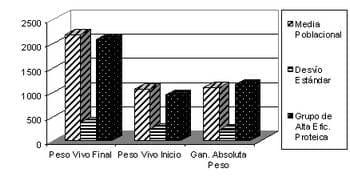
Gráfico 3: Peso vivo y ganancia de peso para la media poblacional y para los 20 pollos con mayor eficiencia proteica entre los 49 (Inicio) y los 79 (Fin) días de vida (gr) (Melo, et al., 2004).
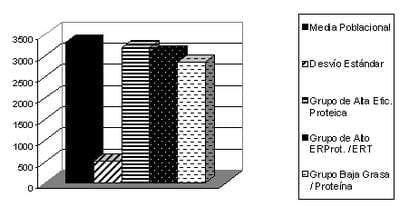
Gráfico 4: Consumo voluntario para la media poblacional y para los pollos con mayor eficiencia en la retención de tejidos entre los 49 (Inicio) y los 79 (Fin) días de vida (gr) (Melo, et al., 2004).
La tasa de retención de proteínas y grasas, como asi también su distribución, depende de variables propias del animal (peso, ganancia de peso, edad, sexo, biotipo, estirpe) como de variables externas (alimentación, clima, manejo, estado sanitario). De acuerdo a Webster (1989), si las condiciones externas no son limitantes, el organismo tiene como objetivo acumular en el tiempo una cantidad determinada de proteína tisular, que determina el tope biológico de crecimiento. Este límite se considera determinado genéticamente. El mismo autor (Webster, 1989) también señala que las variables que afectan la respuesta animal lo hacen en mayor magnitud sobre la cantidad de grasa que sobre la de tejido magro.
La composición química corporal, excluyendo las plumas, cambia a medida que el animal crece. Los cambios mas importantes son el aumento del contenido de grasa y la disminución de agua. La composición de la fracción seca libre de grasas se mantiene constante con el crecimiento del ave (Gous, 1998). En promedio la composición química de un pollo comercial (incluyendo plumas) a los 40 días es: 65 % agua, 19 % proteína, 13 % lípidos y 3 % cenizas.
La mayoría de la materia seca de los músculos, que constituye tan solo un 25% del peso húmedo, se encuentra en forma de proteína (Soller y Eitan, 1984). Este tejido contribuye con el 51 % de la proteína total del ave, mientras que el tejido conectivo lo hace con el 23 %, el hígado con el 5%, el plasma con el 3% y el tracto gastrointestinal con el 5% (Klasing, 1993). La síntesis y degradación proteica son procesos energéticamente caros, siendo baja la eficiencia de su deposición. Es decir, que una gran cantidad de proteína se está depositando mientras otro tanto se esta degradando. Pollos seleccionados por una conversión eficiente de la energía dietaria en proteína corporal lograron altas tasas de deposición proteica con bajas tasas de degradación, en comparación con una población control (Klasing et al., 1987).
El tejido adiposo tiene una cantidad de grasa, agua y una distribución que depende de la dieta, del peso, de la edad y de la raza, biotipo o estirpe. Su composición varía entre un 70 a un 75 % de grasa, 20 a 25 % de agua y 6 a 7 % de proteína, estando distribuida principalmente en tres regiones: piel y subcutáneo (45 %), panículo abdominal (23 %) y entre las fibras musculares (31 %), de acuerdo a las determinaciones realizadas por Carden et al.(1981). El coeficiente de variación para grasa abdominal se encuentra entre un 25 a un 30 %, en comparación con el 15 a 20 % para grasa total, y en total contraposición con los coeficientes de variación para agua, proteína y cenizas (aproximadamente un 2, 3 y 8 %, respectivamente) (Pym, 1988).
Eficiencia Nutricional:
Metabolismo Energético: La energía que se necesita para la síntesis de tejidos y para cualquier otro proceso metabólico que precise energía, es el resultado de la oxidación continua de substratos orgánicos que aportan tanto el alimento, por medio de sus nutrientes, como los tejidos, que se degradan continuamente. Por lo tanto, si la alimentación es insuficiente para mantener al animal, aumenta la degradación tisular y produce una pérdida de peso en el mismo (Di Marco, 1993). La demanda de energía depende del gasto que se realice en los distintos tejidos, lo cual está influenciado por las funciones metabólicas que lleven a cabo los mismos (Webster, 1989). Las funciones metabólicas que demandan energía se agrupan en tres categorías: trabajo fisiológico, transporte de iones de Na+/K+ y biosíntesis de proteínas y grasas. Refiriéndose la primera, por ejemplo, a la actividad del corazón, pulmones y peristaltismo intestinal, y la segunda, a la actividad de las bombas de Na+ y de K+ para mantener el potencial de membrana. En general, dentro del total de energía demandada, la mayor proporción demandada la da el transporte de iones, luego la síntesis protéica, y por último el trabajo fisiológico. En cada tejido predomina alguna de estas funciones, y ello determina las distintas demandas energéticas de los tejidos. Aquellos que cumplen funciones de servicio para el resto del organismo ( cardíaco, pulmonar, hepático, gastrointestinal, renal ) se los ha denominado "supletorios", mientras que los que principalmente estan orientados a una funcion de biosintesis ( muscular, graso, plumas), se los ha denominado "demandantes" (Susbilla et al., 1994). Los primeros, que representan un bajo porcentaje del peso vivo, tienen una alta proporción del flujo sanguíneo y una alta intensidad metabólica, es decir, una importante tasa de síntesis y degradación con un importante gasto en el transporte de iones; siendo el gasto energético por unidad de peso 45 veces mayor en el riñón que en el tejido muscular (Di Marco, 1993). La mayor proporción de peso de órganos digestivos, en parte explica la mayor demanda energética por unidad de peso de los animales jóvenes y de las estirpes genéticas no seleccionadas por eficiencia nutricional y ganancia de peso (Melo et al., 1999; Miquel et al., 1998). Es decir, que a mayor peso disminuye el gasto energético por unidad de peso, lo cual esta contemplado por el concepto de peso metabólico ( Kg0.75 ).
Partición de la energía: Clasicamente, a la Energía Metabolizable (EM), se la puede descomponer en Energía de Mantenimiento (MA), Incremento calórico de Mantenimiento (ICm), Energía Retenida (ER) e Incremento Calórico de Producción (ICp). Se acepta que solo se pueden incluir los términos ER e ICp cuando hay excedente de EM, que entonces se puede destinar a la producción (síntesis de proteína y grasa). También se puede particionar a la energía desde el punto de vista de las funciones metabólicas, ya que es metodológicamente imposible poder discernir entre los procesos de mantenimiento y producción. Los modelos que integran aspectos metabólicos con la respuesta animal utilizan esta partición y calculan el costo de mantenimiento en el proceso iterativo como el punto en que el balance energético es nulo (Di Marco, 1993). La definición clásica de mantenimiento dice que el animal esta en ese estado cuando no gana ni pierde peso, lo cual, desde el punto de vista fisiológico no estaría reflejando casos en los cuales el tejido adiposo es reemplazado por agua, por ejemplo. El mantenimiento es un costo energético variable afectado por la composición corporal y por la proporción de tejidos metabólicamente activos del animal.
Formas de expresar la eficiencia:
Eficiencia energética de la síntesis: Es la energía utilizada en la síntesis de tejidos en relación a la energía total demandada por el organismo. Siendo la formación de tejidos, en general, una pequeña fracción de la ya pequeña que se utiliza en la síntesis de macromoléculas (15-20 %), de acuerdo a Webster (1989). Los tejidos de los órganos internos y vísceras son los principales responsables del gasto de energía a nivel tisular, mientras que a nivel celular lo es el transporte de iones. De acuerdo al mismo autor estas son las principales causas de diferencias en eficiencia entre individuos o biotipos. En términos fisiológicos, se sintetiza tejido mas eficientemente cuanto menor es el peso de los tejidos de alta intensidad metabólica y cuanto menor es el gasto en el transporte de iones, ya que de esa manera hay un menor gasto de mantenimiento (Di Marco, 1993).
Eficiencia en la retención de tejidos: Se asume que las ganancias de peso, tanto del tejido magro como del graso, están directamente relacionadas con un balance energético positivo. Por ello, para que haya crecimiento tiene que haber un exceso de consumo de energía, lo cual determina que el consumo sea uno de los principales causales de la retención energética. Sin embargo, no tiene el mismo costo energético la deposición de un gramo de tejido graso que la de un gramo de tejido magro, ya que si bien demanda la misma energía depositar un gramo de proteína que de grasa, el tejido graso tiene un 20 % de agua y el magro un 80 %, lo cual lleva a que se depositen 0,4 gr y 0,1 gr de tejido magro y graso, respectivamente, por cada kcal de EM (Webster, 1980). Esto determina que a mayor deposición de tejido magro mas eficiente será el individuo.
Eficiencia en la utilización de la EM: Depende del destino de la energía, mantenimiento o producción, y del tipo de tejido que se produzca. El calculo de esta eficiencia asume que el metabolismo de ayuno y el costo de mantenimiento permanecen constantes en cualquier nivel de consumo. No existe duda de que algunos genotipos utilizan la EM mas eficientemente que otros, pero todo indicaría que las diferencias genéticas serían determinadas no por diferente utilización de la energía de producción sino por la de mantenimiento. Estas diferencias se podrían dar por alguno o varios de los factores que se involucran en el mantenimiento del ave, siendo cuatro de los mismos de particular interés: la actividad física, el mantenimiento de la temperatura corporal, el mantenimiento de los tejidos corporales y la tasa metabólica basal (Gous, 1999).
Eficiencia en la conversión de alimento (kg de alimento consumido / kg de peso ganado): Es la mas importante desde el punto de vista productivo. A mayor consumo mayor eficiencia de conversión, ya que se diluye el costo de mantenimiento, por lo cual, los principales determinantes de la eficiencia de conversión van a ser el nivel de consumo, el costo de mantenimiento y el tipo de tejido retenido.
Consumo residual: Debido a que el alimento consumido es utilizado tanto para mantenimiento como para ganancia de peso, se podría afirmar que la Tasa de Conversión es una medida 'bruta' y no 'neta' de la eficiencia. Por otro lado, debido a que el incremento del peso corporal por unidad de alimento consumido es una función decreciente del peso vivo, la ganancia de peso debería corregir las mediciones de consumo a una misma edad (Chambers y Lin, 1988). Los modelos de regresión que incluyen el peso vivo como covariable son usualmente utilizados para ajustar el consumo por diferencias en peso vivo. Esta metodología realiza el ajuste removiendo, al menos, el efecto de las diferencias en el peso. Chambers y Lin (1988) hallaron que la variación remanente luego de corregir el consumo por peso vivo y por ganancia de peso estuvo entre 2 y 9 %, demostrando que las diferencias en consumo entre individuos se debían en gran medida a la tasa de crecimiento. A este remanente de cada medición de consumo con respecto a lo predicho en función de su peso y tasa de crecimiento se lo define como Consumo Residual. Las posibles razones para explicar la variación remanente serían, por un lado, las diferencias entre animales para composición corporal, mantenimiento u otro proceso que demande energía, y, por otro lado, los errores en la predicción y en la medición del consumo (Kennedy et al, 1993).
Medición de la eficiencia en la retención diferencial de tejido
La medición del metabolismo energético y nitrogenado utilizando jaulas individuales no calorimétricas ha sido descripta por varios autores. Parte de los mismos utilizaron como técnica la faena comparativa (Alleman et al., 2000; Geraert et al., 1988; 1990; 1993; Lippens et al., 2002; Melo, 2002; Melo et al., 2004), mientras que otros utilizaron como técnica la recolección de heces individuales (Pym & Farell, 1977; Sklan & Plavnik, 2002). La técnica de faena comparativa precisa la faena previa de una muestra representativa de animales para obtener una estimación de la composición corporal inicial de los animales que continuarán el ensayo. También se obtiene la medición del consumo individual y del peso vivo.
Al finalizar el período de prueba se sacrifican todos los animales que se encontraban en las jaulas. A cada pollo sacrificado se lo despluma y pasa por una picadora hasta homogeneizar el producto final. Del picado final de cada pollo se toman muestras para determinar en cada una de ellas la materia seca, el extracto etéreo y la proteína cruda, de acuerdo a métodos de análisis descriptos por la A.O.A.C. en 1980. La diferencia entre la composición química estimada para el inicio y la obtenida al final del ensayo se utiliza para estimar la grasa y proteína retenida por cada individuo.
La cantidad de energía retenida como proteína y grasa se estimó multiplicando a los gramos retenidos por 5,66 y 9,37 Kcal, respectivamente. Siendo la energía retenida total la sumatoria de las energías retenidas como proteína y como grasa. También es factible estimar la eficiencia proteica debido a que se cuenta con la medición de la cantidad de proteína consumida y retenida (Ret./Cons.).
Resultados obtenidos en distintos ensayos
Los valores medios de la fracción de energía retenida como proteína y como grasa que se obtuvieron varían de acuerdo a la dieta, edad, estirpe y temperatura que se utilizaron para la realización de los ensayos. El incremento proteico en la dieta bajo dietas isoenergéticas no siempre significó un aumento de la media de fracción de energía retenida como proteína, aunque en una línea magra bajo condiciones de termoneutralidad siempre hubo una respuesta positiva al agregado de proteína a la dieta hasta un 23 % (Geraert et al., 1990; 1993). Como era de esperar el aumento de la edad influye negativamente en la fracción de energía retenida como proteína (Geraert et al., 1988), al igual que una temperatura constante de 32 ºC (Geraert et al., 1993), mientras que temperaturas bajas (10 ºC) no dieron resultados distintos a los obtenidos bajo termoneutralidad (Geraert et al., 1988). Estirpes genéticas seleccionados por mayor o menor grasa abdominal dieron como resultado una variación consecuente (Geraert et al., 1988, 1990, 1993), mientras que Jorgensen et al. (1990) encontraron diferencias significativas entre líneas seleccionadas por conversión y por ganancia de peso, siendo esta última de menor proporción de energía depositada como proteína. En estirpes no seleccionadas, Melo et al.(2004) encontraron resultados intermedios a los de las líneas grasa y magra en condiciones de elevada temperatura (Geraert et al., 1993), lo cual era esperable ya que la primera evaluación fue llevada a cabo en verano.
Los valores medios de eficiencia proteica también se ven afectados por una elevada temperatura ambiental, mientras que la disminución a 10 ºC no llevaría a diferencias con la termoneutralidad, de acuerdo a lo informado por Cahaner y Leenstra (1992), Geraert et al. (1988, 1993) y Melo et al.(2004). Como era de esperar la edad también afecta negativamente a la eficiencia proteica (Geraert et al., 1988), mientras que las diferencias entre genotipos son las mismas que las señaladas para la fracción de energía retenida como proteína. Donde se observa una gran diferencia con respecto a la mencionada fracción es en la disminución de la eficiencia proteica con el incremento de proteina cruda en la dieta en un rango entre 13 y 25 % (Geraert et al., 1990, 1993; Sklan & Plavnik, 2002).
Parámetros productivos y otras medidas de eficiencia
En trabajos previos se ha demostrado que las diferencias en el contenido de grasa de dos estirpes genéticas seleccionadas en forma divergente por proporción de grasa abdominal no se pudieron explicar enteramente por diferencias en el consumo o pérdida de energía (Leclercq y Saadoun, 1982; Geraert et al., 1988; Geraert et al., 1990; Geraert et al.., 1993). Sin embargo, si se encontraron diferencias significativas en la fracción de energía retenida como proteína o como grasa, siendo aumentada la fracción retenida como proteína y disminuida la fracción retenida como grasa en la estirpe magra, y viceversa. Melo et al.(2004) encontraron una mayor variabilidad para la fracción proteica (ERP/ERT) que para la fracción grasa (ERG/ERT), lo cual parecería indicar una contraposición con la habitualmente mayor variabilidad de las proporciones de tejidos grasos con respecto a los magros y también en contraposición a la mayor variabilidad de los porcentajes de extracto etéreo con respecto a los porcentajes de proteína bruta corporal. Sin embargo, ambas variables se comportan complementariamente y si se busca a los individuos de mayor proporción de energía retenida como proteína van a ser los mismos individuos que tienen la menor proporción de energía retenida como grasa, lo cual no cambiaría la decisión que se tome con respecto a diferentes genotipos o dietas.
A pesar de existir escasa bibliografía referente a las correlaciones entre las variables de eficiencia en la retención diferencial de tejido y las variables de producción, si se realizaron análisis de varianza para comparar a las líneas magra y grasa mencionadas en el párrafo anterior para estos dos conjuntos de variables. Como ya habíamos señalado, estas líneas se diferenciaron en forma significativa (p<0,05) en las variables de eficiencia en la retención diferencial de tejido, lo cual nos da una noción de la magnitud en que se asociaron con algunas de las variables de producción. Melo et al. (2004) hallaron una correlación cercana a cero entre el peso vivo de pollos de 49 días de una estirpe de crecimiento lento y las fracciones de energía retenida, en concordancia con la ausencia de diferencias significativas (p>0,05) entre los pesos de las líneas magra y grasa encontrados por Geraert et al. (1988;1990) para las 7 semanas de vida. Sin embargo, los citados trabajos muestran que a mayor edad el peso vivo si parecería tener algún tipo de asociación con la fracción de energía retenida, ya que de acuerdo con las correlaciones y coeficientes de regresión obtenidos los animales de mayor fracción proteica retenida fueron de menor peso vivo a los 79 días que los animales de mayor fracción grasa retenida, en animales de crecimiento lento (Melo, et al., 2004), mientras que Geraert et al. (1993) en contraposición a estos resultados informa un mayor peso vivo de la línea magra con respecto a la grasa a los 63 días. En ese trabajo las líneas se diferencian en forma significativa por una mayor ganancia de peso para la línea magra, siendo la misma de mayor ganancia de proteína a pesar de tener un menor consumo de proteína. Esta diferencia deja claro que la mayor ganancia de peso de un individuo no significaría mayor ganancia de proteína en todos los genotipos y condiciones (Gráfico 1).
Gráfico 1: Ganancia de peso (gr) total y por componente químico para los 20 pollos con mayor energía retenida como proteína (ERP/ERT) y los 20 con mayor energía retenida como grasa (ERG/ERT) entre los 49 y los 79 días de vida (Melo et al., 2004).
En la mayoría de los casos el consumo voluntario de los animales se expresó en el mismo sentido que la ganancia de peso, siendo los individuos de menor fracción grasa retenida los que presentaron menor consumo, de acuerdo a las correlaciones y coeficientes de regresión obtenidos por Melo et al.(2004) y en conformidad con lo descrito por Geraert et al.(1988, 1990, 1993) en sus trabajos. Las diferencias en consumo parecerían ser explicadas en gran proporción por el peso vivo y por la ganancia de peso debido a que no hubo diferencias en el consumo residual de estos animales y, por ende, en el consumo corregido por el peso y la ganancia de peso (Gráfico 2). Es decir que la mayor parte del exceso de consumo de los animales de menor fracción proteica retenida se debe a las diferencias en ganancia de peso y peso vivo que a diferencias en su composición química.
El menor consumo de los animales de alta fracción proteica en una población de crecimiento lento no alcanzó para presentarlos como de mejor conversión debido a que también fueron animales de menor ganancia de peso (Melo, et al., 2004). Esto se contradice con lo hallado para las líneas grasa y magra de Geraert et al..(1988,1990,1993), que tuvieron conversiones alta y baja, respectivamente y diferentes en forma significativa (p<0,05).
Eficiencia Proteica: Considerando que las líneas magra y grasa utilizadas por Geraert et al. (1988,1990,1993) también difirieron en eficiencia proteica en forma significativa (p<0,05), se observó que las mismas siempre difirieron en conversión, como resultado de diferencias en consumo y ganancia de peso (Geraert et al., 1993) o solo de Consumo (Geraert et al..., 1990), en concordancia con el sentido de las correlaciones (r=0,47) presentadas por Melo, et al. (2004). La diferencia en eficiencia proteica entre los animales no parecería deberse principalmente a un bajo peso inicial, a pesar que ello determinó la diferencia de ganancias de peso relativa, ya que el consumo residual negativo para los animales de mayor eficiencia proteica podría estar indicando la deposición de una mayor fracción proteica de tejido, debido a que es la mas eficiente de realizar (Soller y Eitan, 1984). Lo cual también se sustenta en que el porcentaje de variación de la eficiencia proteica explicado por el peso vivo a los 49 días que informan Melo, et al., (2004) fue de solo un 9 %, mientras que la proteína bruta corporal lo hizo en un 42 %, de acuerdo al R2 de los modelos de regresión simple respectivos. La diferencia en composición química corporal era preexistente a la medición de la eficiencia proteica y se mantuvo hasta los 79 días, independientemente del peso vivo, ya que a los 49 días había un 45 % de gramos de proteína con respecto al total de gramos retenidos para los 20 animales de mayor eficiencia proteica y 40 % para los 20 pollos de menor eficiencia proteica, diferencia que se mantuvo a favor del primer grupo a los 79 días (Melo, et al.,2004).
Gráfico 2: Consumo (gr) Voluntario y Corregido por peso y ganancia de peso para los 20 pollos con mayor energía retenida como proteína (ERP/ERT) y los 20 con mayor energía retenida como grasa (ERG/ERT) entre los 49 y los 79 días de vida (Melo et al., 2004).
El peso vivo inicial mas bajo para los animales de alta eficiencia proteica podría observarse por la tendencia de los genotipos mas magros a tener una curva de crecimiento mas lenta al inicio, de acuerdo a lo explicitado por Geraert et al.. (1993), en el sentido que los animales magros mostraron menor crecimiento inicial que los grasos pero a los 63 días de vida obtuvieron mayor ganancia de peso, mejor conversión y mayor peso vivo con un menor consumo voluntario (p<0,05). Esto podría estar sugiriendo que los animales de mayor eficiencia proteica serían animales de mayor peso a la madurez, lo cual significaría un mayor requerimiento en los reproductores, pero que al conjunto del sistema le sería mas que compensado por un aumento de eficiencia en el crecimiento de una proporción muy grande de animales en relación a la cantidad de reproductores. Las líneas seleccionadas por Jorgensen et al... (1990) y por Leenstra y Pit (1987) para la mejora de la conversión obtuvieron una mejora correlacionada de la eficiencia proteica (Cahaner y Leenstra,1992), en concordancia con las correlaciones y el signo de los coeficientes de regresión que obtuvieron Melo, et al. (2004).
Discusión
Pym (1996) obtuvo el mismo resultado en conversión para dos estirpes que diferían en un 33 % en grasa, y que habían sido seleccionadas por menor grasa una y por mayor ganancia de peso la otra. En ese sentido, parecería que las variables de producción no tendrían claros efectos sobre las variables de eficiencia en la retención diferencial de tejido. La existencia de correlación genética entre la proporción grasa y la conversión, que no es alta pero es negativa, explica la obtención de respuestas correlacionadas (Jorgensen et al..., 1990; Pym y Solvyns, 1979; Geraert et al.,1990,1993). Sin embargo, existirían otras posibilidades para mejorar la eficiencia nutricional en las cuales no estaría comprendido el cambio en el tipo de tejido retenido, como sería el aumento de la cantidad de energía metabolizable que se puede obtener de la dieta (Jorgensen et al. 1990) o el aumento del turn-over proteico, o la disminución del requerimiento de energía para el mantenimiento, lo cual parecería sugerir que sería necesaria alguna medición de composición corporal directa si se quiere mejorar la eficiencia nutricional por medio de la eficiencia en la retención diferencial de tejido, si fuera el caso donde la composición corporal es parte del beneficio económico. Los bajos coeficientes de determinación obtenidos para la predicción de alguno de los parámetros de eficiencia en la retención diferencial de tejido por medio de variables de producción, aunque se utilizaran conjuntamente (Melo, et al., 2004), parecería indicar una mediana o baja asociación entre los mismos, lo cual sustentaría lo arriba señalado. Esto, sin embargo, no impediría hacer algunas observaciones sobre como se podrían utilizar las variables de producción si se pretende mejorar la eficiencia nutricional por medio de la mejora en la eficiencia en la retención diferencial de tejido. De los resultados obtenidos parecería que existen variables de eficiencia en la retención diferencial de tejido que en forma directa no darían bajos pesos finales, ya que los animales de alta eficiencia proteica son de bajo peso inicial y alcanzan el peso promedio al llegar la edad de faena, obteniendo una ganancia de peso por encima de la media (Grafico 3).
Esto plantearía un crecimiento lento al inicio y aumentar rápidamente en la última etapa previa a la faena, como fuera ya sugerido por varios autores (Jones & Farell, 1992; Leeson & Zubair, 1997; Lippens et al., 2002). Este tipo de curva de crecimiento permitiría un menor gasto de mantenimiento en los animales y una menor deposición de grasa relativa al total de energía retenida, sin dejar de resignar el aumento en la variable que mayor incidencia tiene en el beneficio económico del productor, como es el peso vivo final.
En todos los casos los animales de mayor eficiencia del trabajo realizado por Melo, et al. (2004) tuvieron un consumo voluntario inferior a la media (Grafico 4). Para agregar sustento a lo escrito hasta aquí, Webster (1989), en cierta medida, también recomienda que se considere a mediciones como la eficiencia proteica o la fracción ERP/ERT para la determinación de individuos mas eficientes en la retención de tejidos pero que no resignen peso final, ya que considera que la mejor forma de hacerlo es considerar a la energía retenida como proteína en función del consumo como criterio para lograrlo, lo cual permitiría elegir a los animales de mayor ganancia de tejido magro con el menor consumo posible.
Gráfico 3: Peso vivo y ganancia de peso para la media poblacional y para los 20 pollos con mayor eficiencia proteica entre los 49 (Inicio) y los 79 (Fin) días de vida (gr) (Melo, et al., 2004).
Gráfico 4: Consumo voluntario para la media poblacional y para los pollos con mayor eficiencia en la retención de tejidos entre los 49 (Inicio) y los 79 (Fin) días de vida (gr) (Melo, et al., 2004).
Referencias Bibliográficas
1. Allemann,F, Michel, J., Chagneau,A., Leclercq,B(2000)The effect of dietary protein independent of essential aa on growth and body composition in genetically lean and fat chickens.Br.Ptry.Sc.,41:214-8
2. Assoc. Official Analytical Chemists(1980)Official Methods of Analysis,13thed.AOAC, Washington.
3. Cahaner,A. y Leenstra,F. (1992) Effects of high tº on growth and efficiency of male and female broilers from lines selected for high weight gain, favorable FCR, and high or low fat content. Ptry Sc.,71:1237-50.
4. Chambers,J. y Lin,C. (1988) Age-constant versus weight-constant feed consumption and efficiency in broiler chickens. Poultry Science, 67:565-576.
5. Carden,A., Goenaga,P. y Schang,M. (1981) Efectos de sexo y raza sobre la composición corporal en pollos parrilleros. II. Distribución del músculo y de la grasa. Producción Animal, 7:363-370.
6. Di Marco,O. (1993) Crecimiento y respuesta animal. Ed. Asoc.Arg.Prod.Anim. Balcarce.
7. Geraert,P., Macleod,M. y Leclercq,B. (1988) Energy metabolism in genetically fat and lean chickens: diet and cold-induced thermogenesis. Journal of Nutrition, 118:1232-1239.
8. Geraert,P., Macleod,M., Larbier,M., Leclercq,B.(1990) N metabolism in genetically fat and lean chickens. Poultry Science, 69 :1911-1921.
9. Geraert,P., Guillaumin,S. y Leclerq,B. (1993) Are genetically lean broilers more resistant to hot climate?. British Poultry Science, 34 :643-653.
10. Gous,R. (1998) Growth and development. In: The poultry production guide. Ed. Millet International
11. Gous,R. (1999) Genotype-environmental interactions in laying hens. Memorias de la Jornada de Nutrición y Genética en Aves de Producción, Buenos Aires, 31-42.
12. Jones,G.. y Farrell,D.(1992)Early-life food restriction of broiler chickens I. Methods of application, amino acid supplementation and the age at which restrictions should commence. Br. Poultry Science,33:579-587.
13. Jorgensen,H., Sorensen,P. y Eggum,B. (1990) Protein and energy metabolism in broiler chickens selected for either body weight gain or feed utilization. British Poultry Science, 31: 517-524.
14. Kennedy,B., Werf,J. van der y Meuwissen,T. (1993) Genetic and statistical properties of residual feed intake. Journal of Animal Science, 71:3239-3250.
15. Klasing,K., Jarrell,V. y Calvert,C. (1987) Protein synthesis and degradation in muscles from fast and slow-growing chickens. Poultry Science, 66: 1189-1196.
16. Klasing,K. (1993) Nutritional regulation of protein accretion in chickens. Proceedings of the Symposium Novus Internacional, Méjico, 63-70.
17. Leclercq,B. y Saadoun,A. (1982) Selecting broilers for high or low abdominal fat: Comparison of energy metabolism of the lean and fat lines. Poultry Science, 61:1799-1803.
18. Leenstra,F. y Pit,R. (1987) A comparison among lines selected for less abdominal fat, lower feed conversion ratio, and higher body weight after restricted and ad-libitum feeding. Poultry Sc.,66:193-292.
19. Leeson,S. y Zubair,A. (1997) Nutrition of the broiler chicken around the period of compensatory growth. Poultry Science,76:992-999.
20. Lippens,M., Huyghebaert,G. y De Groote,G.(2002) The efficiency of nitrogen retention during compensatory growth of food-restricted broilers. British Poultry Science, 43:669-676.
21. Melo,J., Miquel,M.C., Mallo,G., Ciacciariello,M. y Villar,E. (1999) Effects of dietary crude protein on slaughter yield of selected broiler stocks. Journal of Applied Genetics, 40(3):219-231.
22. Melo,J.(2002)Evaluación criterios de productividad en pollos. Tesis Maestría Salud Animal FCVUBA
23. Melo,J., Porteyro Ibarra, J., Erias,A; Morao, L.; Cortes Rondan,F.; Miquel,M.C.(2004)Relationship between food efficiency and production traits in a free-range broiler stock. Proc.22nd WPC Ed. Dig.
24. Miquel, M.C., Melo, J, Mallo, G., Villar, E. Castillo, J.L., Ciacciariello, M. (1998) Growth and abdominal fat of “Free range” and commercial broilers. Proc. 6th WCGALP, Armidale, 24: 333-336.
25. Pym,R. y Farrell,D. (1977) A comparison of the energy and nitrogen metabolism of broilers selected for increased growth rate, food consumption and conversion of food to gain. Br. Poultry Science,18:411-426.
26. Pym,R. y Solvyns,J. (1979) Selection for food conversion in broilers :body composition of birds selected for increased body weight gain, food consumption and food conversion ratio.Br.Poultry Science,20:87-97.
27. Pym,R. (1988) Selection for lean tissue growth in broiler chickens. Final Report for Project UQ15C.
28. Pym,R. (1996) Selection for feed efficiency and increased lean tissue growth rate in meat chickens. Proceedings of the XX World’s Poultry Congress,1 :469-478.
29. Sklan,D. y Plavnik,I.(2002) Interactions between dietary crude protein and essential amino acid intake on performance in broilers. British Poultry Science, 442-449.
30. Soller,M. y Eitan,Y. (1984) Why does selection for liveweight gain increase fat deposition? a model. World's Poultry Science Journal, 40:5-9.
31. Susbilla,J., Frankel,T., Parkinson,G., Gow,C. (1994) Weight of internal organs and carcass yield of early food restricted broilers. British Poultry Science, 35: 677-685.
32. Webster,A. (1980) The energetic efficiency of growth. Livestock production Science, 7:243-252.
33. Webster,A. (1989) Bioenergetics, bioengineering and growth. Animal Production, 48:249-269.
Temas relacionados:
Autores:

Universidad Nacional de Lujan
Recomendar
Comentar
Compartir

Universidad Nacional de Lujan
18 de abril de 2007
Estimado Dennis,
En el último Congreso Latinoamericano de Avicultura los investigadores del INTA de Argentina (Marcelo Schang y Jorge Azcona) hicieron una presentación respecto al uso de extracto de alcachofa en dietas para pollos parrilleros, donde se informaba una mayor proporción de lípidos digestibles, una mayor EMA y una mejor conversión de los animales cuando se incluía el extracto de alcachofa en el alimento de 0-21 días, por lo cual remito a los interesados a las memorias del Congreso para tener mayor detalle de los beneficios en eficiencia nutricional.
Estimado Javier,
Si vas a vender pollos por peso vivo en principio parecería que no te tendría que interesar cuál es el tejido que se deposita para lograrlo, pero siempre hay que considerar que cuesta 4 veces más depositar 1 g de grasa que 1 g de tejido magro, por lo cual tendrías que controlar su paniculo de grasa abdominal con cierta periodicidad para lograr una dieta que cubra los requerimientos de los animales pero que no tampoco exceda por mucho los mismos.
Un cordial saludo,
Julián
Recomendar
Responder

Alberta Pork
11 de abril de 2007
Para el Dr Melo: mis felicitaciones. Dr., tengo una pregunta. Para un productor pequeño ¿cómo lo puede ayudar su artículo? ¿qué debería hacer en su finca si produce pollito en bajas cantidadades?
Gracias DR.
Recomendar
Responder
Universidad Nacional de Lujan
4 de abril de 2007
La EMV utilizada en nuestro ensayo fue de 3.400 Kcal aprox., y toda la información con respecto a este ensayo está en las memorias del último Congreso Mundial de Avicultura, llevado a cabo en Estambul.
Recomendar
Responder
Universidad Nacional de Lujan
5 de enero de 2007
En nuestro trabajo presentado en el WPC 2004 no se analizaron diferencias entre animales de diferente sexo, debido a que preferimos disminuir efectos fijos para el análisis, y aumentar el número de repeticiones para obtener las asociaciones descriptas en el trabajo. De todos modos, estoy seguro que la diferencia sería importante, pero que variaría de acuerdo al genotipo que se estuviera evaluando.
Cordiales saludos,
Julián
Recomendar
Responder


7 de junio de 2009
Saludos:
Es un articulo muy completo, lo felicito Dr. Melo, en este estudio presentado se pueden ver muchas variables y muy buenos resultados, es importante mencionar que este tipo de investigaciones no son fáciles de realizar y que son desde cierto punto de vista muy complejos.
Sin embargo los resultados y los detalles de la investigación son muy entendibles y claros, sin duda el articulo está muy completo y es muy interesante por lo menos para mí que soy estudiante y me gusta el campo de la nutrición es muy bueno.
Este trabajo es muy educativo, investigativo, sería bueno evaluarlo por medio de distintas raciones y distintas razas así como también en distintos niveles de producción o periodos de el desarrollo de los pollos y por qué no evaluarlo en otras especies.
Recomendar
Responder

6 de junio de 2009
Ing cual seria el porcentaje de cloruro de sodio para adicionar a la racion de pollos parrilleros.? estoy haciendo un trabajo sobre el tema con los sig porcentajes 0.4[percent], 0.6[percent], 0.8[percent], 1.0[percent] gracias.!!!
Recomendar
Responder
5 de mayo de 2007
Estimado Javier, te recomiendo que revises sobre coccidiosis, ya que si estás mezclando ingredientes sin adicionar un anticoccidial me temo que el riesgo de desarrollo de una coccidiosis es muy alto.
Recomendar
Responder
19 de abril de 2007
Buenos dias, Dr Julian: mi pregunta es ¿qué o con qué mezclo mi alimento para darles mayor consistencia? Ya que me lo están desperdiciando mucho porque queda muy harina, es maíz o sorgo, girasol, semillas de onoto, verduras desidratas molidas, otros residuos de cosecha y un poco de alimemto concentrado comercial en polvo también...Todo esto bien mezclado pero muy polvo. Gracias.
Recomendar
Responder

15 de abril de 2007
Hola: buenas tardes, soy productor de pollos de engorde y estoy ensayando con una alimentación llamada 50-50, que no es otra cosa que suministrarle al pollito en su primera semana el alimento concentrado que me suministra una casa comercial, y luego mezclo un porcentaje de sorgo, uno de maíz, harina de girasol, y materia verde (forraje molido) y he obtenido muy buenos resultados.
Esto sacándolos al sacrificio a las 7 semanas, en un peso de 2 kls, es la tercera ves que lo hago y los comentario de las personas a las cuales les vendo ya beneficiado, me dicen que es más blando, de mejor color y olor, e incluso me comentan que sabe como criollo. Al principio los vendía normales, empecé com 3000, y luego para la segumda sacada ya tenía pedidos y más nuevos clientes, luego en esta tercera, trabajé con 15000 y hasta un frigorífico de la zona que escuchó el comentario me llamó y me ofertó comprarme la producción... Le vendí la mitad para poder cumplir con el resto de los clientes. El dia 08-04-2007 metí el cuarto lote de 18000, los costo los he mejorado comprando puro desecho de cosecha que hay en la zona, ya que ésta es agrícola, pero me gustaría saber qué otro material podría utilizar para mejorar la consistencia del alimento y pelatizarlo, ya que tengo que estar pendiente de ponerle sacos abajo de los comederos por la cantidad de alimento que desperdician y volverlo a reutilizar. Gracias
Recomendar
Responder

9 de abril de 2007
Dr. Melo: realmente espectacular su artículo, eminentemente técnico, y enseña mucho.
Para mejorar la eficiencia en la conversíón alimenticia en pollos de Engorde, en Honduras estamos usando algunos ingredientes naturales como la Alcachofa, ¿cuál es su experiencia al respecto?
Recomendar
Responder
.jpg&w=3840&q=75)
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.




Te puede interesar
.jpg&w=3840&q=75)
Norel Animal Nutrition