Vacas lecheras de alta producción adaptación metabólica durante el período peripartal
Publicado: 7 de abril de 2017
Por: Dr.Ms PhD. Leonardo De Luca –Ex Profesor titular de la Cátedra de Producción de Leche de la U. N. L. Z. Profesor Honorifico U.N L.Z
Introducción
A medida en que los rodeos lecheros aumentan en cantidad y en producción individual, el manejo reproductivo se hace más difícil.
La eficiencia reproductiva del rodeo lechero sigue siendo uno de los aspectos más importantes de la ganadería destinada a la producción de leche. La necesidad de incrementar el tiempo en que la vaca se encuentre alrededor del pico de lactancia requiere de un período entre partos lo más cercano a los doce meses. Debido a que la duración de la gestación es invariable, es importante que las vacas se preñen lo más pronto posible después del parto para lograr mayor productividad. La ineficiencia reproductiva sigue siendo uno de los problemas más frecuentes y significativos de la producción lechera.
Existe una gran tendencia por parte de productores y veterinarios de resolver los problemas de ineficiencia reproductiva por medio de nuevas técnicas de inducción, y sincronización de celos, de Inseminación a tiempo fijo, etc., utilizando diferentes tipos de agentes farmacológicos, cometiéndose el error de no analizar el manejo integral del establecimiento. (Ver Figura 1)
Al incrementarse la producción de leche pasan a primer término las patologías metabólicas a nivel del útero, los ovarios y la mama. Por esta razón, estas afecciones se pueden agrupar bajo la denominación de “enfermedades de la producción”. Se considera como causa etiológica de estas enfermedades, un desequilibrio entre los nutrientes ingeridos con los alimentos, y los aprovechados y/o eliminados.
Dicho desequilibrio provoca el “síndrome de parto” de la vaca de alto rendimiento, entendiendo como tal todos los procesos patológicos que están en relación con la tensión a que se ve sometido el animal, desde la gestación avanzada, hasta el parto y la posterior producción láctea hasta la nueva concepción.
Este síndrome abarca enfermedades con síntomas muy diferentes, que para el clínico se presentan como cuadros independientes, tales como la paresia post-parto, la endometritis o la mastitis, con sus manifestaciones y terapéutica correspondientes. Será primordial la búsqueda del déficit nutricional que originó el problema.
Se observa en la Figura N°1, los cambios más importantes que ocurren en el período peripartal que tienen relación estrecha con desequilibrios nutricionales.
FIGURA N°1

Al final de la etapa gestacional el desarrollo fetal exige aproximadamente: 0,92 Mcal. de Energía, 135 gramos de Proteína, 11 gramos de Calcio, 6 gramos de Fósforo y 0,4 gramos de Magnesio por día. Pero estos requerimientos metabólicos se elevan en la producción del calostro, siendo muy superiores a los demandados por el feto.
Solo para producir 15 litros de calostro desde el día del parto la vaca necesita: 17 Mcal de Energía, 220 gramos de proteínas, 35 gramos de Calcio, 14 gramos de Fósforo y 1,7 gramos de Magnesio, los cuales son ingeridos por medio de la dieta, o son secuestrados desde las reservas corporales para ser transportados a la glándula mamaria.
Específicamente, en términos metabólicos, una vaca que produce 35 litros de leche a los 30 días de lactancia sintetiza por lo menos 4,3 Kilos de lactosa, 1,28 Kilos de proteína y 1,4 Kilos de grasa.(Ver Figura 2)
Esta alta y súbita demanda de nutrientes, la cual está determinada por la creciente actividad del tejido mamario no siempre puede ser satisfecha, y si así sucede, el resultado es el desarrollo de enfermedades metabólicas, siendo la Hipocalcemia Puerperal y el complejo Cetosis – Hígado Graso las patologías más frecuentes.
El reparto de los nutrientes para las diversas funciones fisiológicas tiene distintas prioridades. Las funciones de mantenimiento o lactación tienen prioridad sobre las reproductivas. Por lo tanto, pequeños desajustes nutricionales mostrarán antes sus consecuencias sobre la reproducción, que sobre la producción de leche.
Las causas más frecuentes del fallo reproductivo en la vaca lechera son la endometritis y la mortalidad embrionaria temprana (cerca del 75% del total de las causas), así asociada por los veterinarios y casi nunca por los productores, a un error nutricional.
Incluso las metritis, que tienen un claro componente infeccioso, pueden ser consecuencia de una nutrición inadecuada. Por ejemplo, una deficiencia de calcio iónico (que reduce la motilidad uterina), o un exceso de proteína que induzca un aumento de la concentración de amoníaco en sangre, deprimiendo la actividad de los linfocitos, pueden originar esta enfermedad.
Un reciente estudio británico (Darwash et al., 1999) ha descrito un descenso del índice de fertilidad desde un 65% en el periodo desde el año 1975 al 1982; a un 44% en el periodo desde el año 1995 al 1999, lo que representa un descenso de aproximadamente un 1% anual acumulativo.
Como este descenso de la fertilidad se ha unido a un aumento de la producción lechera, los animales seleccionados genéticamente parecerían ser más eficientes reproductores que no seleccionados. Ante esta observación, varios autores (Ferguson, 1988; Zarate, 1996) han sugerido que la selección genética para obtener vacas de mayor producción ha resultado en una selección concomitante de un ganado con menor capacidad reproductiva (cuadro 1).
Cuadro 1.- Relación entre el nivel de producción yla fertilidad (Ferguson, 1988)
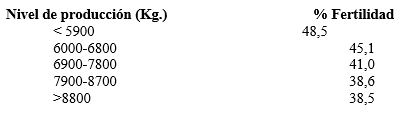
Sin embargo, esta teoría se pone en duda al analizar la capacidad reproductiva de las vaquillonas, que se mantiene equiparable entre los animales no seleccionados y los seleccionados genéticamente. También, en vacas con baja producción láctea y buena fertilidad, la administración de la hormona de crecimiento ha derivado en aumentos de la producción láctea, pero acompañados de un descenso de la fertilidad (Freeman y Lindberg, 1993).
Vaquillonas Holstein con índices de concepción cercanos al 70% demuestran que la raza es genéticamente fértil, aunque sus madres de alta producción láctea presenten índices de alrededor del 40%, lo que confirma que el problema no es genético sino alimenticio.
Existen varias diferencias entre los parámetros reproductivos de las vaquillonas y las vacas lactantes. Estas suelen tener un celo más corto, un ciclo estral más largo, una mayor incidencia gemelar y menor concentración de estradiol en sangre que las vaquillonas. Muchas de estas diferencias han sido asociadas al estado nutricional del animal. Analizaremos cuales son los mecanismos que afectan el funcionamiento reproductivo.
ADAPTACIÓN DEL METABOLISMO HEPÁTICO DURANTE EL PERÍODO DE TRANSICIÓN
El hígado es el órgano de mayor actividad metabólica en los rumiantes, utilizando el 26% aproximadamente del consumo de oxigeno corporal total, mientras que solo representa el 2% del peso total del cuerpo.
Debido a que los nutrientes absorbidos por el tracto gastrointestinal deben pasar obligadamente por el hígado, éste tiene una enorme influencia sobre la calidad y tipo de nutriente suministrado a los tejidos periféricos para el mantenimiento y las funciones productivas y reproductivas.
Durante la etapa de transición el hígado sufre modificaciones funcionales que derivan en cambios metabólicos en las vacas productoras de leche.
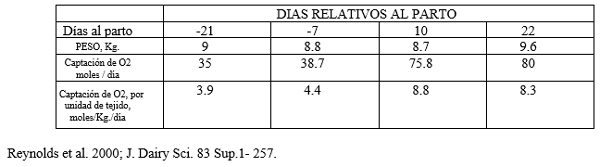
Los datos suministrados en cuadro Nº 2 indican que el tamaño del hígado no tiene modificaciones notables durante el período de transición; sin embargo, la captación de oxígeno como indicador de la actividad metabólica casi se duplica durante la lactación temprana, si se lo compara con la etapa preparto.
CUADRO Nº 2
Este incremento de la actividad metabólica es consecuencia de que el hígado aumenta la síntesis de glucosa después del parto. Calculando el suministro de glucosa proveniente de la absorción intestinal y de la síntesis hepática de los nutrientes absorbidos, éste es 500 gramos menor del total de glucosa que una vaca necesita los primeros 21 días de lactancia.
Esta diferencia se debe suplir con la movilización de las proteínas o sea por la neoglucogénesis de los aminoácidos, durante esta etapa temprana de lactación.
La carga metabólica del hígado aumenta muy rápidamente después del parto, de 4,4 mMoles de O2/g de tejido hepático al día 11 preparto a 8,6 mMoles/g al día 11 post parto lo mismo que el flujo sanguíneo a hígado Se determinó que el que el flujo sanguíneo aumentó 84% desde el preparto (1140 L/h) al postparto (2099 L/h), mientras que el consumo de materia seca aumentó sólo en un 44%, señalando que el aumento del flujo a hígado es una respuesta y no una causa de la mayor actividad metabólica.
Debido a que el consumo de materia seca no es capaz de sustentar el aumento en la demanda de nutrientes para la producción de leche, el aumento en el aporte de sustratos sólo explica una parte del aumento de la actividad metabólica.
La oxidación de glucosa en todo el organismo disminuye después del parto y aumenta la entrada de glucosa, desde el hígado, comparada con el período preparto.
Es posible que la conservación de la glucosa esté dada por el aumento de la concentración de Somatotrofina que ocurre alrededor del parto, puesto que su administración exógena disminuye la oxidación de la glucosa en el organismo.
Sin embargo, el cálculo del aporte de glucosa a partir del consumo de energía digestible subestima la disponibilidad real de glucosa. La salida de glucosa desde el hígado en vacas durante el período de transición se relaciona estrechamente con la demanda de glucosa.
Al día 11 postparto, con una producción de leche de 36 L/día la salida de glucosa hepática alcanza niveles de 2760 g/día y la demanda calculada fue de 2729 g/día; lo mismo sucede al día 22 postparto, con una producción de glucosa en hígado de 3283 g/día para una producción de leche de 41.9 L/día y los requerimientos estimados fueron de 3121 g/día.
FIGURA N° 2
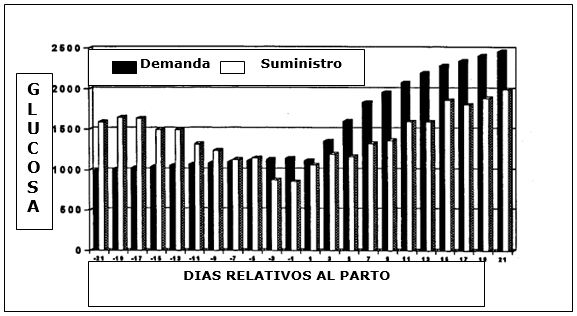
*Demanda de glucosa y aporte durante el período de transición en vacas lecheras. (Piepenbrink y Overton 2000).
Es evidente que luego del parto existe un drástico aumento en la demanda de glucosa. Se estima que el total de la glucosa suministrada por la ingesta energética digestible excede la demanda durante la preñez tardía, pero es menor a la demanda calculada en 500 gramos por día después del parto.
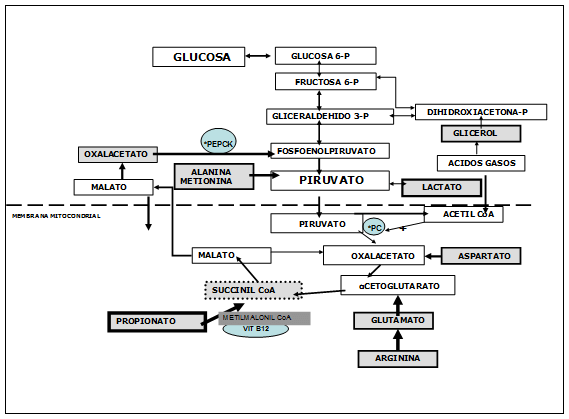
La mayor parte del aumento en los requerimientos de glucosa debe ser obtenido vía gluconeogénesis hepática a partir de los aminoácidos absorbidos en intestino y desde sustratos endógenos como aminoácidos, lactato y glicerol.
La máxima contribución a la gluconeogénesis ha sido estimada en 32 a 73% para propionato, 10 a 30% para aminoácidos, alrededor de 15% para lactato y sólo una pequeña proporción para glicerol. VER FIGURA Nº 3.
Aún cuando no son el sustrato primario, los aminoácidos otorgan una contribución importante a la gluconeogénesis en rumiantes, excepto leucina y lisina, los cuales son completamente cetogénicos.
La alanina y la glutamina contribuyen entre el 40 y 60% del potencial glucogénico de todos los aminoácidos. La contribución de los aminoácidos a glucosa puede ser, por lo menos parcialmente, dependiente de su abastecimiento.
El músculo esquelético es el principal abastecedor de aminoácidos que sustenta el aumento de la gluconeogénesis durante el período postparto.
Los cambios en la capacidad para convertir alanina a glucosa comparada a los cambios en el metabolismo del propionato, sugieren un aumento de la actividad de la enzima piruvato-carboxilasa (PC), que canalizaría cadenas de carbono hacia glucosa desde sustratos que son convertidos a piruvato, incluyendo lactato y los aminoácidos alanina, cisteína, glicina, serina y treonina. VER FIGURA Nº 3
La síntesis de la enzima Piruvico Carboxilasa en hígado aumentó 75 veces al día 1 postparto comparado con valores al día 28 preparto, para luego disminuir a valores mínimos al día 28 postparto.
En contraste, la síntesis de la enzima fosfoenolpiruvato-carboxykinasa (PEPCK) aumentó más lentamente, solamente un 50% al día 28 postparto comparado a los valores preparto, La PEPCK mitocondrial, la cual no responde a la regulación hormonal o al estado fisiológico, es responsable de más del 60% de la formación de glucosa en el hepatocito de rumiantes.
FIGURA Nº 3 PARTICIPACION DEL CICLO TRICARBOXILICO EN TRANSAMINACION Y NEOGLUCOGENESIS
La movilización de la grasa corporal en esta etapa transicional eleva los AGNE (Ácidos Grasos No Esterificados) en el plasma. El hígado los incorpora gradualmente pero una vez saturado, no tiene suficiente capacidad para distribuirlos, por lo que procede a quemar el excedente para proveer energía, o resintetizar lípidos y reencausarlos al torrente circulatorio como Lipoproteínas de Muy Baja Densidad (LMBD). (Ver Figura 4)
Cuando la deficiencia energética post parto es importante se produce una exagerada liberación de los AGNE desde los depósitos, el hígado los acumula y deposita como triglicéridos, conduciendo esto a diversos estadios de hígado graso.
FIGURA Nº 4 bis
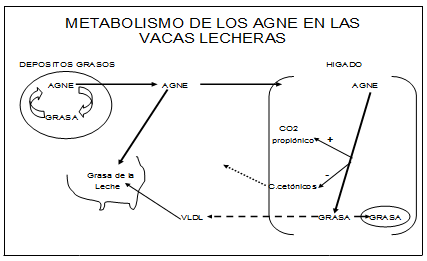
Una importante cantidad de triglicéridos se acumulan en el hígado de las vacas durante el periodo de transición, siendo mas elevada los primeros 21 días post parto. Lo importante es saber en que momento o cual es el límite que el acumulo de grasa es nocivo para la salud de las vacas.
Con un grado de infiltración superior al 15% las funciones ureagénicas (síntesis de urea hepática) se resienten, pero con 30% o mas del parénquima infiltrado, comienza a resentirse la capacidad neoglucogénica propiónica, que es la síntesis de glucosa a partir del propionato ruminal. Si la lipidosis es masiva, la capacidad neoglucogénica a partir de los aminoácidos comienza a ser ineficiente.
Si con solo un 15% del parénquima lipidótico se altera la síntesis de urea proveniente del amonio ruminal, se debe afirmar que una dieta hiperproteica en la fase transicional temprana, es perniciosa para la salud y la producción de las vacas lecheras.
La concentración de amoniaco en la sangre periférica se duplica cuando la infiltración grasa es muy elevada en el post parto inmediato. Los parámetros hemáticos que se alteran en la fase de transición post parto cuando existe una importante lipidosis son las enzimas hepáticas, tales como la GGT y GOT, además de la Bilirrubina indirecta; si las tres se encuentran elevadas se puede confirmar un daño del hígado de tipo distrófico con lipidosis.
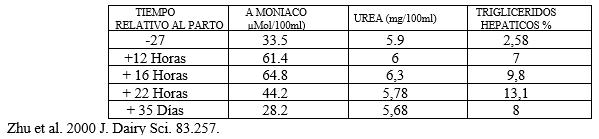
En estas condiciones en el 90% de los casos se encuentra muy elevado el amoniaco sérico con hipoalbuminemia absoluta. Zhu y col. demostraron que la infiltración del hígado con triglicéridos neoformados disminuye la capacidad tanto neoglucogénica como ureagénica.
CUADRO Nº 3 CONCENTRACION DE UREA Y AMONÍACO EN RELACION A LA INFILTRACION GRASA EN EL PERIODO DE TRANSICION DE LAS VACAS LECHERAS
CUADRO Nº 4 CONCENTRACION DE UREA AMONIACO Y ENZIMAS HEPATICAS DURANTE EL PERIODO DE TRANSICION EN VACAS LECHERAS CON PATOLOGIAS PODALES (PROMEDIO SOBRE 386 VACAS ANALIZADAS)
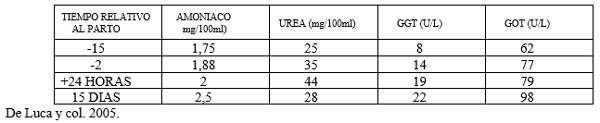
En el Cuadro Nº 4 se puede observar el importante estado de hepatopatía con elevación de los tenores de amoníaco sérico. La disminución de la concentración de Urea sérica va seguida de una elevación significativa del amoníaco, esto demuestra la imposibilidad por parte del hepatocito de captar el amoníaco plasmático.
VER FIGURA Nº 5 - VACA EN FASE TERMINAL DE COMA HEPATICO
VER FIGURA Nº 6 - NECROPSIA DE LA VACA ANTERIOR
VER FIGURA Nº 7 - HIGADO GRASO.
Estrategia para manejar los ácidos grasos no esterificados en las vacas en transición
El manejo de los AGNE en el período de transición es un factor importante que influencia la salud del hígado, la capacidad del mismo para sintetizar glucosa y en consecuencia, su incidencia sobre la producción y los desórdenes metabólicos de las vacas en transición. Los dos principales enfoques que se deben tomar en cuenta son:
- Disminuir el suministro de AGNE hacia el hígado a través de dietas o manejos en la alimentación. Se recomienda incluir en la dieta suplementos neoglucogénicos.
- Optimizar la capacidad del hígado, para que le permita quemar el exceso de AGNE para proveer combustible (energía), o exportarlo hacia el torrente circulatorio como triglicéridos de muy baja densidad (LMBD).
Un buen programa para las vacas a parir y recién paridas, combina un manejo adecuado del medio ambiente y de la alimentación para alcanzar altos niveles de consumo de materia seca (MS). El mejor suplemento que se puede utilizar en la estrategia nutricional prepartal es el suministro de granos termoprocesados tales como Copos de Maíz o Cebada. Con ello se logra elevar la glucemia y disminuir tanto los AGNE como el Acido Beta Hidroxibutírico (el principal cuerpo cetónico encontrado en la sangre).
La segunda estrategia se puede implementar utilizando un suplemento vitamínico que esté capacitado para mejorar la utilización de los Ácidos Grasos No Esterificados. La Carnitina es un elemento necesario para transportar los AGNE dentro de las mitocondrias para que sean quemados. La Carnitina puede ser suministrada a través de la dieta o ser sintetizada por la vaca a partir de la Metionina y la Lisina, aminoácidos reconocidos como limitantes en la síntesis de leche de vaca. Con esto se mejora positivamente la utilización de los AGNE evitando su depósito.
Otra posibilidad que puede ser combinada con la anterior es la de mejorar la exportación de los triglicéridos neoformados elevando la síntesis de Apolipoproteina por el hígado. Para ello se puede suministrar en la etapa de transición el Cloruro de Colina suministrada en forma “protegida” para evitar la destrucción a nivel ruminal, a la dosis de 70 gr. por día por vaca; o la Fosfatidilcolina por vía parenteral.
CICLO DE LA UREA CON RELACIÓN AL METABOLISMO ENERGÉTICO Y PROTEICO
La ureagénesis compite con la gluconeogénesis por el Oxalacetato, un sustrato para el ciclo de Krebbs.
Cuando la capacidad de reacción de los sistemas enzimáticos del hígado son excedidos, como por ejemplo, la saturación de la enzima Carbamoil Fosfato Sintetasa 1, el exceso de amonio debe ser retenido como Glutamato o Glutamina. Este proceso representa un costo para el organismo en cuanto a la disponibilidad de precursores para neoglucogénesis, dado que para la formación de Glutamato se gasta α-Cetoglutarato. Esta es precisamente la reacción que hace del ión amonio un compuesto tóxico en el cerebro.
El cerebro es un órgano muy sensible a niveles altos de amonio y depende casi exclusivamente de la degradación de la glucosa hasta la formación de Acetil-CoA, y su posterior ingreso en el ciclo de Krebbs para formar la energía que requiere. Para que la Acetil-CoA sea metabolizada en dicho ciclo es necesaria una fuente de Oxalacetato, y en el cerebro, esta demanda es cubierta por la carboxilación del piruvato. La enzima que cataliza esta reacción, la Piruvato carboxilasa, es limitada en el cerebro, de tal manera que para el adecuado funcionamiento del ciclo de Krebbs en este órgano es necesario un suministro constante de Oxalacetato.
Cuando se eleva la concentración de amonio serico, en las vacas que consumen dietas ricas en proteínas degradables, se pone en marcha un mecanismo inmediato de detoxificación, la formación de Glutamato a partir del Alfa Cetoglutarato, reacción catalizada por un enzima de alta afinidad la Glutamato sintetasa. Este aminoácido a su vez en presencia de más amonio se transforma en Glutamina, la cual trasporta este anión hasta el riñón donde es eliminado. Por lo tanto este aminoácido permite excretar rápidamente sin pasar por el ciclo de la urea en el hígado, a un metabolito tan tóxico como el amoniaco.
Este proceso al utilizar un intermediario del Ciclo de Krebs como es el Alfa Cetoglutarato hace que los niveles de Oxalacetato disminuyan. De esta manera se reduce drásticamente la síntesis de ATP a nivel cerebral., conduciendo a la aparición de los síntomas de intoxicación amoniacal.
Además es importante tener en cuenta que en un estado de acidosis metabólica, lo cual es frecuente en el postparto de las vacas de elevada producción, se reduce la formación de la urea, y se incrementa la de glutamina. Esto tiene que ver con la generación de ácido carbónico (bicarbonato) que participa en la síntesis de Carbamoil Fosfato, compuesto necesario para que el amoniaco entre al hígado para formar Urea. Por lo tanto a mayor cantidad de amonio que ingresa al hígado, mayor utilización de bicarbonato agravándose de esta manera el estado acidótico.
Por otro lado está establecido que la glutamina es el principal aminoácido involucrado en la amoniogénesis renal, un proceso íntimamente relacionado con la excreción de ácidos. Bajo condiciones de acidosis metabólica, el organismo trata de equilibrar el desbalance entre aniones y cationes a nivel renal mediante la excreción de N en forma de amonio. Es habitual que bajo condiciones de acidosis metabólica el organismo recurra a la movilización de reservas proteicas para garantizar la excreción de amonio y así resolver el equilibrio ácido–base. Definitivamente el metabolismo de los aminoácidos está afectado por el balance ácido-base de la vaca.
Otra característica metabólica de suma importancia para la economía del nitrógeno en los rumiantes, tiene que ver con la necesidad de un segundo átomo de nitrógeno proveniente del aspartato, en el ciclo de la urea.
Al aumentar la utilización de éste aminoácido y exceder la disponibilidad del mismo se estimula su síntesis por transaminación desde otros aminoácidos, con el subsecuente desgaste de las fuentes proteicas que estaban disponibles para ser utilizados en la síntesis de proteínas lácteas,
La detoxificación de amonio para formar urea estimula el catabolismo de la metionina, lo que implicaría que rumiantes alimentados con dietas con alto contenido de proteínas degradables en el rúmen, en el periodo peripartal podrían presentar limitaciones para cubrir los requerimientos de este aminoácido.
El ciclo de la urea se encuentra ligado estrechamente con la neoglucogénesis a través del ciclo de Krebs. Este es un aspecto crítico para el metabolismo de los rumiantes,
Como la absorción de la glucosa es muy baja a nivel intestinal, ocasionado por un disminuido flujo de almidones desde el rumen hacia el duodeno, los requerimientos para este metabolito son muy elevados, particularmente al inicio de la lactancia.
Se debe tener en cuenta que para la síntesis de lactosa el principal componente que determina el volumen de leche producida es la glucosa.
La glucosa, también es fundamental en la síntesis de ácidos grasos y de triglicéridos en la glándula mamaria aportando glicerol, NADPH por la vía pentosa fosfato, y por último ATP. Asimismo participa en la síntesis de proteína aportando esqueletos carbonados para la biosíntesis de aminoácidos no esenciales y ATP.
La glucosa es además esencial en el metabolismo energético a nivel del sistema nervioso y participa en todos los procesos metabólicos de los lípidos. Por su baja absorción intestinal, los rumiantes se consideran animales neoglucogénicos. Esto hace que el incremento en la gluconeogénesis a partir de los aminoácidos, aumente la síntesis de urea.
Además la gluconeogénesis eleva el nivel de nitrógeno amoniacal por la desaminación oxidativa, aumentado el estrés metabólico en los animales de alta producción.
La alanina y especialmente la glutamina (glutamato) son los principales aminoácidos que participan en este proceso aportando átomos de carbono. La contribución de estos aminoácidos representa un importante porcentaje de la glucosa catabolizada (utilizada) y de esta manera pueden suministrar buena parte de los requerimientos netos de glucosa fundamentalmente durante los períodos de bajo consumo de alimentos. Ambos aminoácidos son liberados en gran proporción de las masas musculares del tren posterior durante condiciones de muy baja ingesta de materia seca.
Durante las primeras semanas del inicio de la lactancia se produce una intensa movilización de lípidos desde las reservas, La elevación de los AGNE que se movilizaron y se trasladan al hígado terminan produciendo hígado graso, y sabemos que la acumulación de grasa en el hepatocito reduce la ureagénesis asociada con una disminución en la expresión de algunas de las enzimas que participan en el ciclo de la urea. Esto implica que en las vacas que presentan una extrema movilización de las reservas grasas el amonio toma la vía del glutamato y la glutamina producida eleva la eliminación del amonio vía renal. (Ver Figura 8)
Por ello hemos establecido que la disminución de la capacidad ureagénica que se presenta en las vacas de alto mérito genético cuando se encuentran en estado de hepatopatía lipidótica es responsable de la alta morbilidad y a veces mortalidad que se presenta en el período peripartal, (De Luca L, y col.,2005). (Ver Figura 9)
Rodeos lecheros alimentados con pasturas de alto tenor proteico, de alta degradabilidad ruminal, y con bajo contenido en hidratos de carbono no estructurales producen poca proteína bacteriana, altos tenores de amonio ruminal y baja utilización del nitrógeno. Estos animales demuestran en los perfiles metabólicos graves alteraciones en los parámetros relacionados al metabolismo energético y proteico, además de presentar importantes aumentos de las enzimas hepáticas.
Un ejemplo clásico es el de un rodeo lechero de 967 vacas en lactancia al cual se le realizo un Perfil Metabólico a un 7% de los animales en producción
La dieta fundamentalmente era pastoril a base de alfalfa en estado de prefloración con una baja y/o errónea ingesta energética. Los animales demostraban diferentes patologías metabólicas, baja producción láctea, baja producción de grasa, y proteína láctea, alto porcentaje de vacas con síndromes de tipo cetósico, parálisis ruminal, aletargamiento, abortos de 3 a 5 meses, 5% de vacas con patología podales sobreagudas, infosura y muertes súbitas. Los análisis en general arrojaron los siguientes resultados promedio: Urea sérica se mantiene baja a muy baja (10-17mg/ dl ± 4), albúmina baja (2,2g/dl ± 0,86), glucemia muy baja (30mg/dl ± 8,68), AGNE elevados (1,25 mg/dl ± 0,22), amoníaco elevado (2,33 mg/dl ± 0,88), GGT elevada (> 22U/L), GOT elevada (>76U/L), Bilirrubina total elevada (1,22 mg/dl).
Esto demuestra que, no solamente se encuentra disminuida la ureagénesis, además las vacas demuestran un grave desbalance en la relación energía /proteína, con un balance energético negativo extremo y acidosis metabólica. (Ver Figura 10)
La sobrecarga metabólica y energética puede contribuir a incrementar el balance energético negativo y alterar la función reproductiva. Con raciones bajas en energía, el catabolismo del exceso de nitrógeno puede representar un factor importante sobre la reproducción.
Existe una reducción del índice de concepción del 85 a 56% al comparar raciones que aportaban el 65% de los requerimientos energéticos y contenían un 15 o un 21% de proteína bruta. Este costo energético debe contrastarse con los beneficios que puede aportar un exceso de nitrógeno, como puede ser un incremento de la ingestión a través de un aumento de la digestibilidad de la ración
El aporte de niveles de proteína adecuados, especialmente en conjunción con un buen perfil de aminoácidos, (proteínas pasantes, grano de soja termo procesado) no sólo puede mejorar el balance energético del animal (reduciendo la síntesis de urea), sino que además puede aumentar la síntesis de lipoproteínas. Las lipoproteínas, especialmente la LDL y mas la HDL son ricas en metionina y lisina.
Aportar niveles suficientes de estos aminoácidos es esencial para asegurar que la síntesis de estas lipoproteínas sea adecuada.
Las lipoproteínas son agentes esenciales para el transporte de colesterol, molécula necesaria para la esteroidogénesis (estrógenos y progestágenos).
Carroll et al. (1992) demostraron que niveles elevados de LDL y fundamentalmente HDL aumentaban la secreción de progesterona por parte del cuerpo lúteo en el vacuno lechero.
Además de participar en los aportes de colesterol, las lipoproteínas participan en la estimulación directa de la secreción de IGF-I por parte de las células luteales
Podemos establecer el balance metabólico de las vacas lecheras cuando las mismas son sometidas a dietas altas en proteínas degradables:
- Un gramo de urea excretada por conversión del amonio cuesta 5,6 Kcal.
- 100 gramos de proteína cruda que no se utiliza ocasiona una pérdida de 0,2 Mcal de Energía.
- Un exceso de 1000 gramos de proteína cruda ocasiona un costo energético para conversión del amonio de 2 Mcal/ día (este gasto representa el 7% del requerimiento energético para producir 30 litros de leche)
- Cuando el Balance Energético Negativo es de -11 Mcal, el gasto representa el 18% del requerimiento energético para producir 30 litros de leche.
- Por ello, en estas condiciones se atrasa la primera ovulación, y cuando las mismas ocurren los cuerpos lúteos formados tienen poca capacidad de síntesis progesterónica.
Los mecanismos que explican los efectos negativos de la alta ingesta proteica sobre la reproducción incluyen.
- Alteración del eje Hipotálamo- Hipófisis- Ovario, directa o indirectamente (por cambio en el metabolismo energético)
- Alteración del metabolismo energético en forma directa influenciando la producción de hormonas o su metabolización o eliminación.
- Efectos tóxicos directos del amonio sobre la viabilidad espermática o embrionaria.
- Alteración de las tasas de absorción de algunos minerales tales como; Calcio, Magnesio, Cobre y Zinc.
Es conocido que la hiperamoniemia condiciona a los animales a una inmunodepresión por inhibición directa de la Linfoblastogénesis, de la síntesis de la diversas Citokinas Linfocíticas y por último el estado Hiperureico altera fuertemente la marginación neutrofílica por deficiencia en la síntesis de L-Selectinas a nivel del endotelio capilar
El estado hiperamoniacal en el inicio de lactancia conduce a diversas alteraciones tanto productivas como reproductivas. Este estado subclínico desencadena cuadros de infertilidad
- La concentración ruminal de MH4 para la óptima función ruminal es de 2,9 mmol/L a 5 mmol/L
- La digestión de la fibra es más eficiente a una concentración de MH4 superior a 2,9 mMol/L e inferior a 5mmol/L (15 mg%)
- Concentraciones mayores a 5.5 mmol/l esta asociado a alcalosis ruminal, lo cual determina, menor contracción muscular del rúmen, disminución de la digestibilidad ruminal, disminución del consumo voluntario.
- Una mayor concentración de Urea/Amonio en sangre conduce a menor producción de progesterona por el cuerpo lúteo, formación de cuerpos lúteos quísticos y frágiles.
- Mayor amonio en sangre elevan el contenido de Zinc en el lumen uterino, bajan los niveles de Fosfatos, Potasio y Magnesio, esto afecta la capacitación espermática, la fertilización y altera la interacción entre las hormonas y el conceptus en la etapa preimplantatoria, (mayor mortalidad embrionaria).
- Mayor desciliación y descamación de la mucosa de la trompa por lo tanto altera la fertilización y el transporte tanto de espermatozoide como el ovulo fecundado.
ESTRATEGIA DE LA SUPLEMENTACIÓN
La suplementación en la vaca seca depende de dos factores, o aspectos nutricionales. El primero, está referido al estado corporal de la vaca (suplementación con granos) y el segundo, a la suplementación con sales aniónicas.
Con respecto al primero, debe ser evaluado en el último mes de lactancia puesto que si se realiza cuando la vaca está seca difícilmente podamos solucionar problemas inherentes al estado corporal.
Se sabe que la vaca es más eficiente para transformar granos en tejidos cuando está lactando, por lo tanto al secado la vaca deberá estar en buen estado corporal. En cuanto al segundo, el período óptimo de suplementación es de 30 días antes del parto, por cuanto al ser las sales de baja palatabilidad, necesita ese período de acostumbramiento.
CONSIDEREMOS LA CONDICION CORPORAL
Los lineamientos comunes sobre calificación corporal óptima se basan en los siguientes factores:
- Las vacas gordas comen menos antes y después del parto y tienden a bajar su ingestión más pronto ya antes de parir. Esta condición está relacionada al síndrome de la vaca gorda, patología muy común en los establecimientos donde los intervalos entre partos son mayores a 14 meses con períodos de seca superiores a los 95-100 días, como así también cuando las dietas en el pre parto son demasiado altas en energía. En estos casos recomendamos inyectar con Selenio y Vitamina E (ambos antiditróficos) y algún protector hepático a base de Metionina, Cisteína, Arginina), y contemplar la posibilidad de agregar a la dieta Colina elemento imprescindible en la profilaxis de la lipidosis.
- El aumento en la calificación de condición corporal de 2 a 3 al parto, aumenta la producción de leche en 320 a 330 litros durante los primeros 90-95 días después del parto. Teniendo una calificación de 4 en vez de 3, aumenta solo de 33 a 40 litros en este período con el peligro de sufrir el síndrome de la Vaca Gorda. Con calificación mayor a 4 no existe aumento de producción, sino una disminución manifiesta con patologías relacionadas al metabolismo lipídico/cálcico (acidosis, desplazamiento abomasal, retenciones placentarias y/o loquiales, coma puerperal).
- La mejor producción de leche ocurre con una calificación corporal entre 3 y 4, independientemente del número de lactancia.
- Mejorar la condición corporal de las vacas flacas (aumento en la escala de 2 a 3) puede elevar la producción de leche hasta en 500 litros en la futura lactancia y por supuesto mejora la eficiencia reproductiva.
- Las vacas que tienen menos de 3 puntos en la calificación corporal entre el parto y la primera inseminación artificial, tienen tasas de concepción de 38%, comparadas con el 50% de las que tienen nivel 3, y con el 65% de las vacas que superan esta calificación.
- La ingestión de materia seca es el concepto más crítico relacionado con la energía. La baja ingestión del período pre-parto, impone la cantidad real de nutrientes a suministrar (kilos de grasa, kilos de fibra, gramos de almidón, y megacaloría de energía). Las vacas requieren kilos de nutrientes no porcentajes. Las vacas que consumen más materia seca cuando están próximas al parto tienen mayor ingestión 21 días después del parto. Todo lo que podamos hacer para mejorar la ingestión de materia seca es ganancia. Revisando algunos parámetros nutricionales vinculados al consumo podemos evaluar situaciones relativas a la ingestión de nutrientes.
- La adición de almidones con alta degradabilidad ruminal puede mejorar y aumentar la síntesis de proteína bacteriana ruminal cuando los forrajes son ricos en proteínas solubles, elevando la tasa de pasaje de nitrógeno a nivel intestinal.
- La inclusión de 1 a 2 kilos de paja de trigo o fibras de baja digestibilidad puede mantener lleno el rumen y permitir su correcto funcionamiento, evitando de esta manera el síndrome de vaciamiento ruminal, factor clave en el desencadenamiento del desplazamiento abomasal.
- Si se somete a las vacas al principio del período seco bajo una dieta restringida, se puede evitar la ganancia excesiva de peso y la reducción posterior en ingestión de alimentos. Así se evita el desencadenamiento del Síndrome del Hígado Graso.
- La alimentación con niveles importantes de ensilado de maíz en las raciones para vacas próximas al parto ayuda a la ingestión, reduce los niveles de potasio en la dieta, y proporciona almidones fermentables para el rumen. Si a esto se le puede adicionar maíz de alta humedad en pequeñas proporciones (1 a 2 kilos) la relación de almidones degradables/altamente degradables, es óptima conduciendo a una excelente transición al período post parto.
- Al usar una ración con características similares a la utilizada en el período de lactancia, nos aseguramos una transición sin trastornos digestivos, como es la acidosis ruminal latente, patología subclínica que precede a la Cetosis y al coma hipocalcemico.
- Debemos tener sumo cuidado cuando adicionamos sales aniónicas con muy baja palatabilidad, esto lleva a bajos consumos de materia seca conduciendo a los animales a graves problemas relacionados a los niveles energéticos.
- Otro de los factores que afectan la ingestión de materia seca es un manejo inadecuado del comedero cuando se incluyen vaquillonas en el grupo, la competencia puede llegar a ser un factor decisivo sobre la ingesta energética.
Por lo tanto las recomendaciones que debemos considerar incluyen:
- Proporcionar energía adicional a las vaquillonas próximas a su primer parto, a las vacas bajo estrés ambiental y a las que tienen más baja condición corporal.
- Desafiar a las vacas próximas al parto que coman más de 14.5 kilos de materia seca (el valor normal es de 11.8 a 12.0 kilos en los últimos 30 días antes del parto). Debemos tratar que las vaquillonas durante este período consuman más de 11.5 kilos (el valor normal es de 10 kilos).
- Proporcionar algo de fibra de partícula larga (cantidades limitadas de heno de buena calidad basándonos en sus niveles de potasio y uso estratégico de paja de trigo o avena)
- Debemos incluir forrajes de buena calidad para evitar dietas altas en fibra detergente neutra.
- En la dieta debemos limitar la cantidad de grasa a no más de 100 a 150 gramos por día la cual puede ser suministrada con la incorporación de 1 kilo de semilla de algodón, esto expone a las vacas a una fuente importante de grasa mejorando la ingesta adicional de energía.
- Debemos optimizar los niveles de almidón de alta digestibilidad tanto a nivel ruminal como intestinal, esto estimula por un lado el crecimiento bacteriano (síntesis proteica) y por otro mejora la oferta de glucosa a nivel del intestino delgado manteniendo de esta manera la glucemia a niveles anticetogénicos. Esto lo podemos lograr con cereales termoprocesados (copos de maíz) o en su defecto con maíz de alta humedad mezclado con maíz seco molido con zaranda de 10 milímetros.
CONSIDEREMOS LA SUPLEMENTACION CON SALES ANIONICAS
Si asumimos que las patologías post parto, fundamentalmente las hipocalcemias comatosas son prácticamente una especie de intoxicación subclínica por potasio (K) por ser este catión un inductor de alcalosis, es razonable deducir que el uso de las llamadas sales aniónicas es la solución a estos problemas.
Como señalamos antes, las dietas pre parto altas en fósforo (más de 80 gramos por día) y bajas en calcio (menos de 40 gramos por día), utilizadas desde los trabajos pioneros de Boda y Cole, dejan de ser eficientes en los sistemas reales de producción actuales donde los niveles de producción son altos, y de ninguna manera podemos reducir en forma sustancial el consumo de forrajes ricos en potasio (K), generalmente henos o silajes de pasturas a base de leguminosas.
Constatar los niveles de potasio del forraje refleja solo una parte del problema, la variación en los aniones, especialmente del cloro (Cl-), también puede tener un gran impacto en la diferencia entre aniones y cationes.
Análisis de los principales cationes (K y Na) y los aniones (Cl y S) en ensilados de maíz, ensilado de alfalfa y de praderas, o sea mezclas de pastos y tréboles.
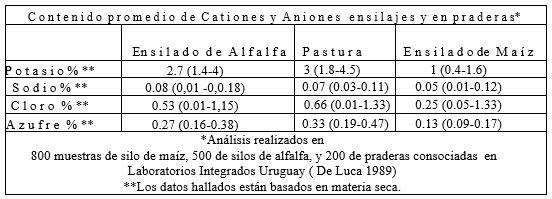
*Análisis realizados en 800 muestras de silo de maíz, 500 de silos de alfalfa, y 200 de praderas consociadas en Laboratorio Integrados Uruguay (De Luca 1989)
**Los datos hallados están basados en materia seca.
Como vemos hay variaciones importantes en los niveles de potasio en el silaje de alfalfa y en la pradera los cuales promedian 2.7 y 3.0 respectivamente, con rangos desde 1.4 a más de 4. Los niveles de sodio son más bajos y no tienen demasiada variación, los de cloro variaron desde 0.01 hasta más de 1.3%, estas variaciones pueden tener un gran impacto en el balance real de aniones y cationes en forrajes y raciones.
Generalmente los niveles de azufre variaron con el contenido de proteína en los forrajes, pues este anión está asociado con los aminoácidos azufrados, por lo tanto encontramos concentraciones que oscilaron entre 0.16 hasta 0.47%.
En el ensilado de maíz también encontramos variaciones importantes, fundamentalmente en el potasio y en el cloro.
Ahora veamos que pasa con las DCAD en los análisis de tres materias primas utilizadas para racionar vacas en transición:
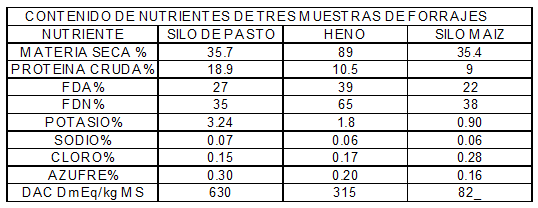
*DCAD =Diferencia Catiónica Aniónica de la Dieta
Se puede observar en este cuadro los resultados de los análisis reales de forrajes de tres muestras, los de silo de pasto y los de silo de maíz, difieren en DCAD por el contenido de potasio y de cloro manteniéndose el sodio constante.
Siendo superior el contenido del anión azufre en el silaje de pasturas, es evidente que la cantidad de potasio genera la peligrosa DCAD en este forraje, seleccionando para el uso en las vacas en transición sin duda los silajes de maíz (+82mEq/Kg. de MS).
El heno si es bajo en potasio (menor de 1.5%) y alto en cloro (superior a 0.19%) es generador de una DCAD menor a 300 mEq/Kg. MS el cual lo transforma en un nutriente apto sin ser peligroso.
Estos resultados nos demuestran la importancia de hacer análisis de forrajes para determinar el contenido de cationes y aniones fijos y poder de esta manera balancear las dietas correctamente. Solo de esta manera estamos absolutamente seguros que la estrategia nutricional para vacas secas en transición está en un camino correcto.
Por todo lo expuesto, debemos convenir que el uso de sales aniónicas queda definido por:
- La cantidad de cationes fijos (especialmente K) que tenga la dieta base.
- La posibilidad y conveniencia de reemplazar a los componentes de la dieta preparto que los contengan.
- El potencial de producción de los animales con los que se trabaja.
- El uso de las sales aniónicas podrá controlar los siguientes problemas:
- Si el ingreso de calcio en el preparto de las vacas de alta producción es de 40 a 45 gr./vaca/día,
- Si la DCAD (Diferencia Catiónica Aniónica de la Dieta) es superior a los +100 a +200 mEq/Kg. MS,
- Si la incidencia de vacas caídas es superior al 3% anual (vacas con más de dos partos),
- Si la mortandad es superior al 20% de los animales caídos.
Una formulación de sales aniónicas que nos ha dado resultados excelentes y que hemos utilizado desde 1990 contiene: Sulfato de magnesio, Sulfato de calcio, Cloruro de amonio, suplementada con Fosfato dicalcico. La cantidad de Cloruro de amonio varía de acuerdo a la cantidad de potasio que contengan los componentes de la dieta preparto, generalmente oscila entre 8% al 16%.
COMO REALIZAR UN MONITOREO DEL ESTADO ÁCIDO-BASE
Una vez que hemos iniciado la alimentación de las vacas secas con dietas especiales utilizando como base a las sales aniónicas debemos determinar si están funcionando correctamente.
Si asumimos que utilizando una dieta que tenga una DCAD baja o negativa simplemente porque realizamos los cálculos sobre un papel estamos equivocados, puesto que no sabemos con seguridad si logramos el objetivo deseado.
Una forma para determinar si funciona es evaluar el número de casos de hipocalcemias comatosas o tetánicas antes y después de haber incluido las sales aniónicas.
Otra técnica es tomar muestras de sangre y realizar análisis de Calcio total y del iónico, puesto que por cada unidad de DCAD que baja, sube una de Calcio total y 1.5 de Calcio iónico.
El método más directo de evaluar la eficiencia de agregar las sales aniónicas es revisar el pH de la orina.
Los cambios en el pH urinario ocurren dentro de los siguientes dos a cuatro días de alimentar con sales aniónicas.
Una pregunta muy frecuente cuando ponemos en práctica un programa de medición de pH en la orina es: ¿Como afecta el horario de la comida al pH urinario?
Cuando las vacas se alimentan varias veces por día no hay efecto del tiempo sobre el cambio de pH, sin embargo cuando se alimentan una sola vez al día, el pH de la orina varía con el tiempo transcurrido desde la hora del consumo del alimento. Con este razonamiento se establece un programa de medición de pH de la orina de la siguiente manera:
- Colectar la orina de 10 vacas como mínimo
- Seleccionar un horario fijo entre las dos a seis horas después de dar de comer
- Tomar muestras en el mismo horario una vez a la semana, el mismo día.
- Usar papel estándar de pH o un medidor digital para uso en el campo. Son muy precisos y económicos.
- Hacer cambios en la ración para alterar la DCAD si el pH promedio de la orina está por fuera del rango deseado de 6.0 a 6.5 en Holstein, o de 5.8 a 6.2 en Yérsey.
Las lecturas mayores a estas recomendaciones indican que la ración no está produciendo los efectos deseados. Se debe constatar los cálculos de la ración suministrada y reformular la dieta.
Si se realizan análisis para determinar el valor de los ingredientes, que sea de química húmeda; el sistema NIR o Cromatografía de Rayos Infrarrojos no es exacto para minerales.
BIBLIOGRAFIA
ADASHI E.Y., HSUEH A.J.W. y YEN S.S.C. (1981) Endocrinology. 108: 1441-1449.
AUBOIRON S., DURAN D., ROBERT J.C., CHAPMAN M.J. y BAUCHART, D. (1995) Repro.Nutr. Dev. 35: 167-178.
BAKER J., HARDY M.P., ZHOU J., BONDY C., LUPU F., BELLVE A.R. y EFSTRA DIADIS A. (1996) Mol. Endocrinol. 10: 603-918.
BAO B., THOMAS M.G. y WILLIAMS G.L. (1997) J. Anim. Sci. 75: 3235-3245.
BAUMAN D.E. y CURRIE W.B. (1980) J. Dairy Sci. 63: 1514-1529.
BEAM S.W. y BUTLER W.R. (1997) Biol. Reprod. 56: 133-142.
BLANCHARD T., FERGUSON J.D., LOVE L., TAKEDA T., HENDERSON B., HASLER J. y CHALUPA W. (1998) Am. J. Vet. Res. 51: 905-908.
BRITT J.H. (1995) Follicular development and fertility: Potential impacts of negative energybalance. Proc. III Congr. Int. Med. Bov. ANEMBE. Santander.
BRUCKENTAL I., DRORI D., KAIM M., LEHRER H. y FOLMAN Y. (1989) Anim. Prod. 48: 319-329.
BUTLER W.R. (1998) J. Dairy Sci. 80: 2533-2539.
BUTLER W.R. y ELROD C.C. (1991) Nutrition and reproduction relationships in dairy cattle. Nutr. Conf. Feed Manuf. Ithaca, NY. pp: 73-82.
CAMPBELL M.H. y MILLER J.K. (1996) J. Dairy Sci. 81: 2693-2699.
CARROLL D.J., GRUMMER R.R. y MAO F.C. (1992) J. Anim. Sci. 70: 2516-2526.
CASSADY J.M. (2000) Initial body composition modulates reproductive response of heifers tonutritional manipulation. M.Sc. Thesis, University of Minnesota.
CHAMBERLAIN D.G., MARTIN P.A. y ROBERSTON S. (1989) En: Recent Advances in AnimalNutrition. pp: 175-193.
CHILLIARD Y., FERLAY A., DELAVAUD C. y BOCQUIER F. (1998) Int. J. Obesity. 22 (Suppl. 3): S171 (Abstract).
COLLINS A., PALMER E., BÉZARD, J., BURKE J., DUCHAMP G. y BUCKLEY T. (1997) Equine Vet. J. Suppl. 25: 12–16.
DARWASH A.O., LAMMING G.E. y WOOLLIAMS J.A. (1999) Anim. Sci. 68: 527-532.
DARWASH A.O. y LAMMING G.E. (1999) Cattle Practice. 7: 397-399.
DASGUPTA P.R., KAR A.B. y DAHR M.L. (1971) Indian J. Exp. Biol. 9: 414-415.
DE LUCA L. J., CASAGRANDE J. (1988) Revista Brasilera de Reproducción. Recife Pernambuco UNP. Recife Brasil. N° 10; 121-144.
DE LUCA L. J., CAROU N. (1987). Desarrollo foliculogénico, síntesis estrogénica, y capacidad ovulatoria en vacas sometidas a régimen de Balance Energético Negativo. Veterinary Practice 12.333-337.
DE LUCA L. J., (1997). Usos de cereales termoprocesados en las dietas de las vacas en transición. Estudios de los parámetros relacionados al BEN del puerperio inmediato. Revista de nutrición zootécnica. 3-123-130- Bologna. Italia. Gianni Guidolin Termofichi.
DE LUCA L., CAPAUL E., GRIMOLDI, R. y SILVA J. 1973. “Método de graficación en perfiles metabólicos” Congreso de Bioquímica Huerta Grande Córdoba. Actas del Congreso. Vol. II.
DE LUCA L., CAPAUL E., GRIMOLDI, R., y SILVA J. - 1973 “Perfiles metabólicos: herramienta fundamental para una mejor explotación del tambo”. Premio de la Academia Nacional de Medicina Veterinaria. Boletín oficial.
DE LUCA L.J., 1976. II Jornadas Latinoamericanas y IV Uruguayas de Buiatría, organizada por el Centro de Médicos Veterinarios de Paysandú. Uruguay. Trabajo presentado. Perfiles Metabólicos en el Bovino de Leche.
DE LUCA L.J., 1976 Primeras Jornadas del Círculo de Médicos Veterinarios de San Luis. Tema. Nutrición y Fertilidad. Miembro disertante
DE LUCA L.J., 1976 VI Jornadas Internacionales de Veterinaria. II 1º Jornadas Veterinarias de Maldonado Uruguay Tema Metabolismo y Fertilidad en la Vaca Lechera. Miembro disertante
DE LUCA L.J., 1976. ”Trastornos metabólicos en la vaca lechera” Sociedad Rural de Trenque Lauquen Pcia de Bs. As. Conferencia para productores y profesionales. En los Anales de la Soc. Rural de Trenque Lauquen.
DE LUCA L.J., 1976 "Perfiles Metabólicos. "Sincronización de celo”. Actualización. Instituto de Ciencias Agronómicas. Universidad Nacional de Córdoba. Conferencia para profesionales y alumnos.
DE LUCA L.J., 1976. “Perfiles metabólicos. Su aplicación a campo”. Sociedad Rural de San Francisco. Córdoba. Conferencia para profesionales y productores. Trabajo publicado en el Anuario de la Soc. Rural de San Francisco.
DE LUCA L.J., 1977. “Perfiles metabólicos. Su aplicación práctica”. Facultad de Ciencias Veterinarias. Esperanza Santa Fe. Conferencia para Profesionales y alumnos. Trabajo publicado por el Centro de Estudiantes de la FCV.
DE LUCA L.J., 1977. Panelista en mesa redonda sobre el tema “Nutrición, Metabolismo y Fertilidad en la vaca lechera” Primera Jornada Veterinaria de Azul. Conferencia para Profesionales.
DE LUCA L.J. ,1978 VI Jornadas de Buiatría Uruguayas. “Eficiencia Reproductiva. Factores limitantes no específicos”. Miembro invitado especial Disertante y panelista. Publicado en los Anales de la Jornada de Buiatría.
DE LUCA L.J., 1980 “Importancia de los minerales en la reproducción”. Primeras Jornadas de Fisiología de la Reproducción en la Vaca Lechera e Inseminación Artificial. Organizado por la Sociedad de Medicina Veterinaria del Uruguay. Atlántida Uruguay. Miembro disertante.
DE LUCA L.J., 1981 V Jornadas Sobre Reproducción Animal Organizadas por CIAVIT Venado Tuerto. Santa Fe. Tema: Aplicación y Valor Práctico de los Perfiles Metabólicos. Miembro disertante
DE LUCA L.J., 1982 IV Congreso Argentino de Ciencias Veterinarias Organizado por Sociedad de Medicina Veterinaria. Tema: Carencias Minerales en los Animales Domésticos. Panelista. Disertante.
DE LUCA L.J., 1986. XIV Jornadas Uruguayas de Buiatría. Paysandú Uruguay. Tema: Perfiles Metabólicos. Algunas Consideraciones. Inducción del Celo, su aplicación Práctica. Miembro disertante.
DE LUCA L.J., Capaul E., 1975 "Fertilidad en el bovino; aplicación práctica de algunos parámetros hemáticos” Actas del Congreso Mundial de Veterinaria Grecia. Tomo II Pág.389.
DE LUCA L.J., Capaul E., 1980 “Retención placentaria no específica; una aproximación profiláctica” Actas del IX Congreso Internacional de Reproducción e Inseminación Animal, Madrid. España Vol. I Pág. 657.
DE LUCA L.J., 1980. “Enfermedades metabólicas del bovino” Sede Facultad de Ciencias Veterinarias. UBA. Directores de Curso Post-Grado Dres. De Luca L.J. y Capaul E., Colaboradores. Dr. González G., y Dra. Carcagno A. Duración tres días.
DE LUCA L.J., CAPAUL E., 1984 ”Retención placentaria un método de prevención” Veterinaria Argentina. Vol. II Pág. 220 a 226.
DE LUCA L.J., CAPAUL E., 1986. “Perfiles metabólicos, algunas consideraciones”. Conferencia realizada en las XIV Jornadas Uruguayas de Buiatría Paysandú.
DE LUCA L.J., 1998. Alimentación y nutrición de la vaca lechera. Curso de actualización. 2 al 30 de Octubre. Universidad Nacional de Lomas de Zamora Facultad de Ciencias Agrarias.
DE VRIES M.J. y VEERKAMP R.F. (2000) J. Dairy Sci. 83: 62-69.
DIRKSEN G.U., LIEBICH H.G. y MAYER E. (1985) Bov. Pract. 20: 116-120.
EDMONSON A.J., LEAN I.J., WEAVER L.D., FARVE T. y WEBSTER G. (1989) J. Dairy Sci. 72: 68-78.
ELROD C.C. y BUTLER W.R. (1993) J. Anim. Sci. 71: 694-701.
ELROD C.C., VAN AMBURGH M. y BUTLER W.R. (1993) J. Anim. Sci. 71: 702-706.
FERGUSON J.D. (1994) J. Dairy Sci. 77: 113-120.
FERGUSON J.D. y CHALUPA W., (1989) J. Dairy Sci. 72: 746-766.
FERGUSON J.D., GALLIGAN D.T., BALNCHARD T. y REEVES M., (1988) JAVMA. 192: 659-662.
FREEMAN A.E. y LINDBERG G.L. (1993) J. Dairy Sci. 76: 3143-3152.
FRICKE P.M. y WILTBANK M.C. (1999) Theriogenology. 52: 1133-1143.
FRIEDMAN J.M. y HALAAS J.L., (1998) Nature. 395: 763-770.
GARCIA BOJALIL C.M., STAPLES C.R., THATCHER W.W. y DROST M. (1994) J.Dairy Sci. 77: 2537-2548.
GILBERT R.O., SHIN S.T., RABUFFO T.S. y CHANDLER S.K., (1996) J. Dairy Sci. 79: 2377-2381.
GLUCKMAN P.D., BREIER B.H. y DAVIS S.R. (1987) J. Dairy Sci. 70: 442-466.
GRIEVE D.A., KOVER S., RIJPKEMA Y.S. y HOF G., (1986) Livest. Prod. Sci. 14: 239-254.
GRUMMER R.R., (1995) J. Anim. Sci. 73: 2820-2833.
GUSTAFSSON A.H. y CARLSSON J. (1993) Livestock Prod. Sci. 37: 91-105.
HARRISON R.O., FORD S.P., YOUNG J.W., CONLEY A.J. y FREEMAN A.E. (1990) J. DairySci. 73: 2749-2758.
HOGGARD N., HUNTER L., TRAYHUN P., WILLIAMS L.M. y MERCER, J.G., (1998) Proc.Nutr. Soc. 57: 421-427.
HOUSEKNECHT K.L., BAILE C.A., MATTERI R.L. y SPURLOCK M.E. (1998) J. Anim. Sci. 76: 1405-1420.
HUHTANEN P, (1998) Agric. Food Sci. 7: 219-250.
HUNTINGTON G.B. (1990) Rep. Nutr. Dev. 30: 35-47.
JI, S.Q., WILLIS, G.M., SCOTT, R.R., y SPURLOCK, M.E. (1997) J. Anim. Sci. (Supl. 1):167 (Abstract).
JORDAN E.R. y SWASON L.V. (1979) J. Anim. Sci. 48: 1154-1158.
JORDAN E.R., CHAPMAN T.E., HOLTAN D.W. y SWASON L.V., (1983) J. Dairy Sci. 66: 1854- 1862.
KAUR H. y ARORA S.P. (1995) Nutr. Res. Rev. 8: 121-136.
KIRKLAND R.M. y GORDON F.J., (2001) J. Dairy Sci. 84: 233-240.
KLUCINSKI W. y TARGOSWKI S.P., (1984) Inmunopharmacology. 8: 47.
KUMAR B., FRANCIS S.M., SUTTIE J.M. y THOMPSON M.P. (1998) Comp. Biochem. Physiol. 120: 543-548.
LADENHEIM R.G., TESONE M. y CHARREU E.H. (1984) Endocrinology. 115: 752-756.
LOEFFLER S.H., DE VRIES M.J. y SCHUKKEN Y.H., (1998) J. Dairy Sci. 81: 14-22.
LUCY M.C., (2000) J. Dairy Sci. 83: 1635-1647.
LUSSIER J.G., MATTON P. y DUFOUR J.J., (1987) J. Reprod. Fert. 81: 301-307.
MANN G.E., MANN S.J. y LAMMING G.E., (1996) J. Reprod. Fert. Abs. 17: Abstract 55.
MAUER R.R. y BEIER H.M., (1976) J. Reprod. Fert. 48: 33-41.
MAYER E., (1986) Bull. Acad. Vet. France. 59: 159-173.
MIHM M., GOOD T.E., IRELAND J.L., KNIGHT P.G. y ROCHE, J.F., (1997) Biol. Reprod. 57: 1328-1337.
NRC (1989) Nutrient Requirements of Dairy Cattle. 6 th Ed. Washington, D. C., Natl. Acad. Press.
NRC (2001) Nutrient Requirements of Dairy Cattle. 7th Ed. Washington, D. C., Natl. Acad. Press.
OLDICK B.S., STAPLES C.R., THATCHER W.W. y GYAWU P., (1997) J. Dairy Sci. 80: 1315- 1328.
OLSON P.A., BRINK D.R., HICKOK D.T., CARLSON M.P., SCHNEIDER N.R., DEUTSCHER G.H., ADAMS D.C., COLBURN D.J. y JOHNSON A.B., (1999) J. Anim. Sci. 77: 522–532
PHERSON B., PLYM K. y CARLSSON J. (1992) J. Vet. Med. 39: 187-192.
RABIEE A.R., LEAN I.J., GOODEN J.M. y MILLER B.G. (1999) J. Dairy Sci. 82: 39-44.
ROCHE, J.F., AUSTIN, E.J., RYAN, M., O’ROURKE, M., MIHM, M. y DISKIN, M.G. (1999) J.Reprod. Fertil. 54: 61-71.
RYAN D.P., BAO B., GRIFFITH M.K. y WILLIAMS G.L. (1995) J. Anim. Sci. 73: 2086 - 2093.
SASSER R.G., WILLIAMS R.J., BULL R.C., RUDER C.A. y FALK D.G., (1988) J. Anim. Sci. 66: 58-63.
SCHRICK F.N., INSKEEP E.K. y BUTTLER R.L., (1993) Biol. Reprod. 49: 617-621.
SHINGFIELD K.J., JOKELA M., KUASTELL K., HUHTANEN P. y NOUSIANINEN J. (1999) Agric. Food Sci. 8: 365-392.
SMITH W.L. y MARNETT L.J. (1991) Biochim. Biophys. Acta 1083: 1.
SPICER L.J., ALPIZAR E. y ECHTEERNKAMP S.E., (1993) J. Anim. Sci. 71: 1232-1241.
SPICER L.J., TUCKER W. y ADAMS G.D., (1990) J. Dairy Sci. 72: 2179-2183.
THACTCHER W.W., STAPLES C.R., DANET-DESNOYERS G., OLDICK B. y SCHMITT E.P., (1994) J. Anim. Sci. 72 Suppl.3): 16.
VALK H. y EBEK L.B.J., (1999) J. Dairy Sci. 82: 2157–2163.
WENNINGER A. y DISTL O., (1994) Deutsche Tierärztliche Wochenschrift. 101: 152-157.
WHERMAN M.E., WELSH T.H. y WILLIAMS G.L., (1991) Biol. Reprod. 45: 514-523.
WILLIAMS J.S., GARDINER C.S., SCHULLER L.S. y MENINO A.R. (1987) J. Anim. Sci. 65 (Suppl. 1): 415 (Abstract).
WILTBANK, M.C., FRICKE, P.M., SANGSRITAVONG, S., SARTORI, R. y GINTHER, O.J. (2000) J. Dairy Sci. 83: 2998-3007.
WIT A.A.C., CESAR M.L.F. y KRUIP T.A.M., (2001) J. Dairy Sci. 84: 1800-1804.
WU Z. y SATTER L.D., (2000) J. Dairy Sci. 83: 1052–1063.
ZHOU Y., XU B.C., BAUMAN G. y KOPCHICK J.J., (1997) Proc. Natl. Acad. Sci. USA. 94: 13215-13220.
FIGURA 1.
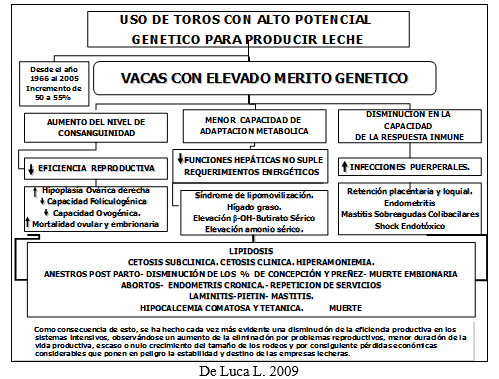
FIGURA 2.

FIGURA 4.
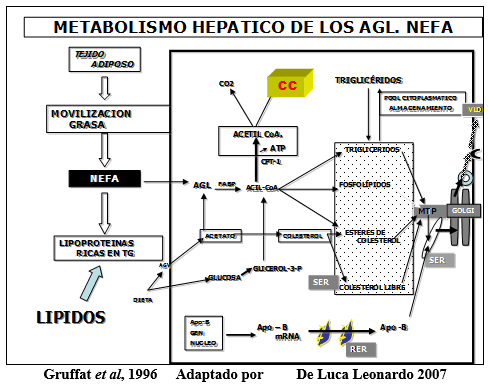
CPT-1 Carnitina-Palmitoil-Tranferasa 1. Enzima regulatoria necesaria para que los Acyl-CoA ingresen a la mitocondria.
Gráfico que explica el destino metabólico de los AGL en el hígado de la vaca en transición.
Los AGL, llegan al hígado y tienen tres destinos.
- Se queman a C02 y H20 + ATP-
- Se reesterifican y forman triglicéridos (grasas)
- Forman cuerpos cetonicos.
Es importante destacar que las vacas en la etapa transicional tienen la poca capacidad de exportar hacia la sangre los lípidos, como LMBD. Para formar esta lipoproteína se necesita un transportador, La Apolipoproteína B, si hay deficiencia de Metionina, Colina, y Carnitina no hay síntesis de Fosfatidilcolina y por lo tanto no se forma este transportador. Esto agrava el hígado graso.
La cetogénesis hepática esta regulada; 1) por el abastecimiento de los AGL al hígado, 2) por la actividad de CPT-1 para promover la entrada de grasa Acyl-CoA hacia la mitocondria para la producción de Acetyl-CoA y 3) por la actividad intramitocondrial de 3-hidroxi-3-metilglutaril-CoA sintetasa (HMG-CoA sintetasa), la cual es el paso regulatorio en la conversión de Acetyl-CoA a cuerpos cetonicos.
Una elevación del ácido propionico desde el rúmen eleva específicamente un intermediario del ciclo del ácido cítrico mitocondrial la Succinil CoA, un potente inhibidor de la HMG-CoA sintetasa. De esta manera bajan los cuerpos cetonicos. Lo mismo pasaría si elevamos la glucemia.
FIGURA 5.

NECROPSIA DE LA VACA ANTERIOR
FIGURA 6
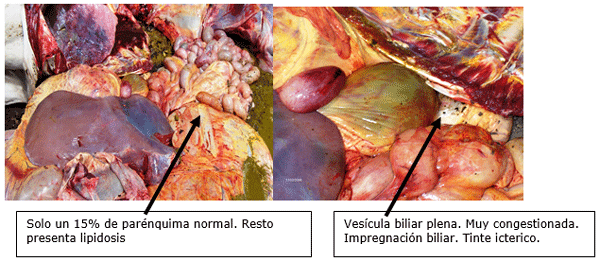
FIGURA 7.
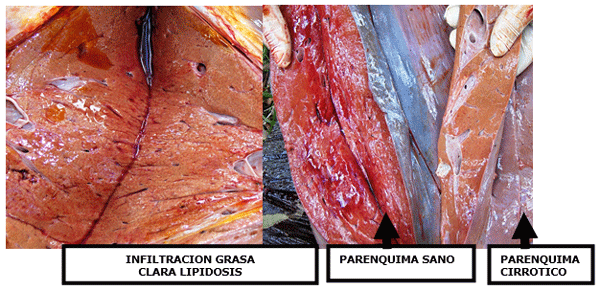
FIGURA 8.
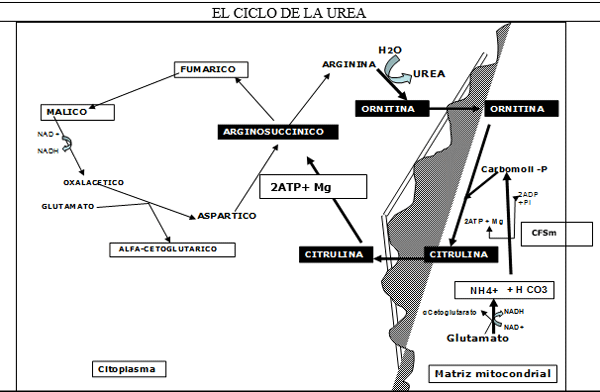
CbP= Carbamoil Fosfato.
CFSmi =Carbamoil Fosfato Sintetasa Mitocondrial. Desde 30% hasta un 80% del Nitrógenos absorbido se realiza en forma de Amonio. Este N2 absorbido puede entra al ciclo como tal, para integrar el CbP. pero la reacción de la CFS para el amonio de baja afinidad. La otra forma es como Glutamato. La Glutámico Dehidrogenasa tiene mucha mayor afinidad para el amonio. Esto tiene mucha importancia ya que cuando hay exceso de amoniaco la enzima CFS se satura. Por lo tanto el amonio se mantiene como glutámico /glutamina.
Si el exceso se mantiene, y la destoxificacion se hace por la formación de Glutamato/Glutamina, el sistema se depriva de intermediarios neoglucogénicos (α-cetoglutarato), representando para el organismo un elevado costo energético, a veces llegando a la falla de síntesis de ATP. (Falla del ciclo tricarboxílico) definitivamente esto conduce a los síntomas neuronales de intoxicación amoniacal. Es importante destacar la importancia que representa el Magnesio en el ciclo de la Urea
FIGURA 9.

FIGURA 10. SINDROME CLINICO DE COMA HEPATICO. FASE INICIA

Temas relacionados:
Autores:

SIPA Servicio Integral de Produccion Animal
Referentes que Recomendaron :
Luis Albornoz, Fernando Hidalgo y Teran Serralde y 3 másRecomendar
Comentar
Compartir

9 de abril de 2017
Muy buen articulo
Recomendar
Responder

9 de abril de 2017
Estoy de acuerdo en el punto que hace el Dr Ponce. Sucede comúnmente en la América Tropical en vacas de tipo lechero Europeo que la nutrición durante el periodo seco se descuida completamente. Aunque los animales poseen el mérito genético a través de anos de IA, el ganadero, continua con los métodos tradicionales de nutrición y no prepara las vacas secas durante las 3-4 semanas antes del parto, tal como Oscar López menciono anteriormente, el resultado final, son animales con una condición corporal muy baja después del parto, y una lenta recuperación de ese peso durante el resto de la lactación.
Esta situacion ha creado un reto para el profesional en esta parte del mundo, en donde los forrajes de buena calidad son difíciles de cultivar y los granos son importados; ambos indispensables en el tratamiento del problema.
Otro fenómeno traído por esta situacion de completa ignorancia de los hechos, es que ganaderías con razas puras y con muy buen material genético, han cometido el error de cruzar el rebaño con otras razas, con la creencia mal fundada de aliviar el problema.
Recomendar
Responder

9 de abril de 2017
Excelente articulo y tambien los demas publicados por al autor. Pocos ganaderos ponen el interes en los trastornos metabolicos asociados a problemas de composicion lactea y aun mas grave en la reproccion. No basta con tener vacas altas productoras sino se gestan pues por ahi se va la plata sin que se le ponga la atencion requerida. Hace tiempos atras investigue bastante en los temas de altercaciones metabolicas en el rumen y el sindrome de leche anormal, algunos de los cuales se han debatido en este forum. En todos esos casos la expresion de los desbalances tambien impactan la reproduccion y y tambien tienen efecto sobre incrementos de la mastitis subclinica. Excelente y ojala todos los ganaderos se lo lean en algunos casos requieren del apoyo de algun tecnico que les ayude a interpretar el articulo y asociarlo a su rebaño, Lo mas importante es saber que hacer en ese periordo critico de la lactancia. Aunque se piensa que estos problemas no existen en el tropico en vacas con baja produccion tambien ocurren pues vienen de un periodo seco con hambre y no han recuperado su condicion corporal y puede que no presente pico de lactancia pero cada problema hay que analizarlo en su contexto. saludos. Pastor
Recomendar
Responder


INIFAP México
8 de abril de 2017
Excelente información...Clara y concisa. Felicitaciones..
Recomendar
Responder

26 de febrero de 2021
Mil Gracias por el aporte profesional y de experiencia vivida saludos de Ecuador
Recomendar
Responder

15 de abril de 2017
felicitaciones un trabajo muy ilustrativo bien redactado y con experiencia, continuen presentando este tipo de trabajos. Abrazos
Recomendar
Responder
13 de abril de 2017
Exelente artículo
Recomendar
Responder
12 de abril de 2017
MAGNIFICO INFORME E INVESTIGACION. COMPRENDO LA MAYOR PARTE DE LA EXPLICACION,
SERIA ESTUPENDO HACERLO EN IDIOMA DE POCA CULTURA DE EMPLEADOS PARA GUIARLES
MEJOR A UNA MAYOR CALIDAD DE CONTROL ALIMENTICIO COMO DE LOS CELOS DE LAS VACAS.
Recomendar
Responder

10 de abril de 2017
Hola totalmente de acuerdo creo que hay muchos productores que no tienen un buen manejo nutricional de sus rodeos.
como ejemplo yo siempre pregunto cuando empieza la lactancia para poder darle la importancia de manejo y nutricion??
para mi la respuesta sienmpre esta empezando lactancia, en el parto pero tambien en prepato pero tambien en seca pero tambien en lote de cola o lote media o lote punta cuando las queremos preñar .
siempre estan empezando lactancia .
Nosotros manegamos la genetica por merito neto. la nutricion esta planteada para todas las categorias igual todo el año sin importar el precio, (si no me alcanza para dar de comer saco vacas pero nunca alimento mal).
y la reproduccion se puede lograr 60% dc y 35 % tc con el manejo correcto.
si todo eso funciona no hay grandes problemas de trastornos. siempre trabajando muchisimo en manejo y confort.
saludos
Recomendar
Responder

10 de abril de 2017
Excelente artículo, a tomar muy en cuenta hoy en que trabajamos con vacas de alta producción cuyos requerimientos de alimentos también exigen ser precisos para no generar descompensaciones, buen material felicitaciones , gracias
Recomendar
Responder


¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar
.jpg&w=3840&q=75)