Efecto del estatus nutricional en la respuesta productiva del ganado criancero: Conceptos básicos para mejorar la eficiencia y rentabilidad.
Publicado: 22 de agosto de 2016
Por: Dr. Rodrigo A. Arias Inostroza, Ing. Agrónomo, PhD, MS in Animal Science.
Introducción
La inminente apertura del mercado chino a las carnes rojas nacionales en conjunto con la coyuntura sudamericana, que incluye el problema sanitario de Paraguay y la importante baja en el número de cabezas bovinas en Argentina, constituyen en sí hechos que replantean la necesidad urgente de desarrollar e implementar una política de aumento de la masa ganadera en Chile. En este sentido, la superficie total de praderas existentes en el país, incluyendo praderas sembradas, mejoradas y naturales, permite alcanzar al menos unos diez millones de cabezas, es decir, prácticamente triplicar la masa existente hoy en día. En el rubro carne bovina, el logro de esta meta requiere de un plan ganadero nacional que incluya entre otros aspectos: manejo de praderas, genética, manejo nutricional, manejo reproductivo, formación de capital humano y la articulación vertical y horizontal de la industria de la carne bovina. Sólo a modo de ejemplo, actualmente existe una gran diversidad de razas, afectando negativamente la homogenización del producto final y complicando el manejo productivo y reproductivo. En este sentido, la articulación con las plantas faenadoras y exportadoras debiera ser mayor, particularmente en lo concerniente a la especificación del tipo de animal que requieren a fin de que los productores se habiliten para entregar un producto estandarizado en términos de peso y calidad de la canal.
Al revisar las estadísticas nacionales en la última década, se observa una clara tendencia a la reducción en la existencia animal. Sin embargo, a partir del año 2007 el precio del ganado en sus distintas categorías ha manifestado un aumento progresivo, el cual se ha acrecentado en los últimos dos años. Como ejemplo, el precio del novillo vivo presentó un incremento del 43,5% en el 2010 respecto del año 2005, según datos del boletín de ODEPA (2011). Situación similar se observa en la categoría terneros(as) en la que se observa un aumento en los precios de aproximadamente un 60% para el último año (diciembre 2009 vs. 2010). En este contexto, debemos recordar que hasta no hace mucho tiempo atrás el negocio de la crianza se consideraba poco rentable y de alto riesgo de capital.
No obstante, la bonanza de precios, la rentabilidad de la crianza se explica en gran medida por la eficiencia reproductiva del rebaño, cuya meta es lograr que cada vaca pueda parir un ternero(a) vivo anualmente. En la práctica, muchos vientres no logran la preñez dentro del periodo de encaste o bien este se extiende más allá de lo recomendado. Una de las principales razones de esto se debe al incremento del intervalo postparto primer estro (IPE), el que se asocia a una inadecuada ingesta de nutrientes. El objetivo del presente artículo apunta a describir el rol de la nutrición en la eficiencia reproductiva de los rebaños crianceros y su aplicabilidad en Chile.
Ciclos estrales en el bovino
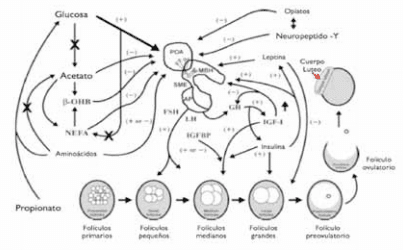
La reanudación del estro, en un plazo relativamente corto después del parto, representa un hito importante para conseguir los objetivos reproductivos del rebaño criancero. Cabe señalar que el órgano que realmente controla los ciclos estrales es el eje reproductivo (hipotálamo–hipófisis–ovario), y no el aparato reproductor como se podría pensar. Este eje es el encargado de integrar las señales nutricionales y del medio ambiente con la respuesta reproductiva. La Figura 1 ilustra las estructuras críticas que participan en el reinicio de la actividad reproductiva después del parto en la vaca, así como los principales mecanismos de regulación. Los centros cerebrales del eje reproductivo están estructurados de tal manera que las neuronas neurosecretoras del área preóptica y del hipotálamo medial basal terminan en el tallo de la eminencia media (Hess y col., 2005). Estas neuronas liberan la hormona liberadora de gonadotrofina (GnRH) al sistema portal hipofisario, el que lo transporta a la glándula pituitaria anterior. Aquí, la GnRH estimula la síntesis y secreción de las gonadotropinas: hormona folículo estimulante (FSH) y hormona luteinizante (LH). Estas hormonas regulan el crecimiento y desarrollo folicular (FSH), y a través de ésto la producción de estrógenos y progesterona en el ovario. Tanto la LH como FSH son secretadas en forma de pulsos y son reguladas por dos sistemas, el tónico y el cíclico (Callejas, 2001). La LH asiste en la maduración final del folículo preovulatorio dominante e influye el desarrollo del cuerpo lúteo. La producción de estradiol por los folículos ováricos eventualmente alcanza un umbral que causa que las neuronas neurosecretoras del área preóptica liberen un pulso de GnRH, lo que a su vez genera una liberación LH (un pulso de gran amplitud) lo que provoca la ovulación. La ovulación genera la formación del cuerpo lúteo, el que produce progesterona, hormona que prepara el revestimiento del útero para recibir el huevo fertilizado y la glándula mamaria para la producción de leche. En caso de preñez se mantiene el cuerpo lúteo, lo que junto al desarrollo de la placenta genera una alta producción de estradiol y progesterona, generando una retroalimentación negativa que causa el anestro durante la preñez y el puerperio debido a la supresión de liberación de GnRH en el hipotálamo (Hess y col., 2005). En caso contrario, la regresión lútea junto a la disminución de la concentración sérica de progesterona permitiría el reinicio del ciclo estral.
La glándula pituitaria almacena y libera las reservas de LH. Cabe señalar que estas reservas son recuperadas de forma relativamente rápida después del parto. Sin embargo, la hipersensibilidad al efecto del estradiol contribuye a la extensión del anestro posparto (Short y col., 1990). Las vacas comprometidas nutricionalmente también parecen ser más sensitivas a la retroalimentación negativa del estradiol (Wettemann y col., 2003) y pueden permanecer sin ciclar por 100 o más días, debido a la disminución en la amplitud y frecuencia de la secreción de LH (Schillo, 1992). No obstante, existe evidencia de que las concentraciones de gonadotrofinas que contiene la glándula pituitaria anterior son similares a las de vacas en estro a los 30 días, y estas responden normalmente a GnRH exógena. Por ello, durante los últimos años la investigación se ha centrado en identificar los mensajeros metabólicos y endocrinos que puedan influenciar los mecanismos que afectan la secreción de LH (Wettemann y col., 2003).
Como se observa en la Figura 1, la glucosa es uno de los substratos más importantes requeridos para el normal funcionamiento del proceso reproductivo. Esto se explica porque este es el principal combustible metabólico utilizado por el sistema nervioso central, y su ausencia reduce la liberación de GnRH (Wettemann y col., 2003). El aumento de los niveles de glucosa en sangre puede lograrse a través del manejo alimenticio que promueva la gluconeogénesis en el animal. Por el contrario, una dismicución en el nivel de glucosa promoverá la movilización de las grasas, la acumulación de ácidos grasos no esterificados (NEFA) y la síntesis de cuerpos cetónicos, aumentando los niveles en plasma de acetato, β-hidroxibutirato y NEFA, lo que conlleva a una reducción en la amplitud del pulso y concentraciones de LH (DiCostanzo y col., 1999). Más detalles respecto del rol de cada factor se puede encontrar en al artículo de Hess y col. (2005).
Figura 1. Mecanismos asociados al estro postparto en vacas de crianza y regulación metabólica. POA= Área preóptica del hipotálamo; MBH= Hipotálamo medial basal; SME= Tallo de la eminencia media; AP= Glándula pituitaria anterior; FSH= Hormona folículo estimulante; LH= Hormona luteínizante; NEFA= Ácidos grasos no esterificados; β-OHB= β-hidroxibutirato; GH= Hormona de crecimiento; IGF-I= Factor de crecimiento insulínico tipo I; IGFBP= Proteína ligante del IGF (Modificado a partir de Hess y col., 2005).
Nutrición y reproducción
Los primeros indicios de la importancia del estatus nutricional en la reproducción datan de principios de los años 40. Hoy en día existe cuantiosa evidencia científica que señala la relación entre la nutrición sobre algunos parámetros reproductivos. En efecto, si el consumo de nutrientes es limitado y la condición corporal es inadecuada tanto vacas como vaquillas cesan la ovulación (Richards y col., 1989; Bossis y col., 1999), aún cuando no se conocen completamente los mecanismos por los cuales el hipotálamo, la pituitaria y el ovario responden al estatus nutricional o a las reservas energéticas (Lents y col., 2005). Según Correa-Orozco y Uribe-Velásquez (2010), lo anterior no está exento de polémica y faltan algunos puntos aún por aclarar. Según Hess y col. (2005), la complejidad de mecanismos biológicos asociados entre nutrición y reproducción pueden ser categorizados en tres grandes áreas: 1) procesos fisiológicos que pueden estar envueltos en la mediación nutricional sobre la reproducción; 2) relaciones entre estatus nutricional y medidas que son indicativas de éxito reproductivo; y 3) ejemplos específicos de manipulación nutricional que inciden en la reproducción. Los mismos autores indican que luego de realizar una revisión de la literatura publicada en el Journal of Animal Science: a) la nutrición en preparto es más importante que la nutrición posparto en la determinación del largo del anestro posparto; b) una ingesta inadecuada de energía durante el periodo final de la preñez reduce la reproducción aun cuando la energía dietaria sea suficiente durante la lactancia; c) una condición corporal (CC) mayor o igual a 5 (escala 1 a 9) asegura que las reservas corporales son adecuadas para la reproducción posparto; y d) reducciones adicionales en la reproducción se producen cuando las vacas se encuentran en balance energético negativo durante el periodo de lactancia.
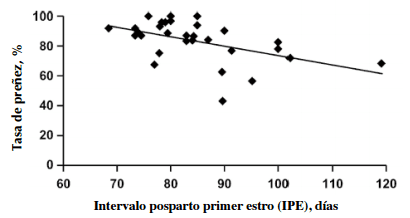
Dunn y Moss (1992) y recientemente, Hess y col. (2005) demostraron que existe una relación inversa entre el intervalo postparto primer estro (IPE) y la tasa de preñez (Figura 2). Asimismo, demostraron que una vaca con un IPE de entre 40 y 60 días tiene un 88% de probabilidad de parir cada 365 días. En este sentido, la relación negativa se explica más bien por los factores que causan la extensión del IPE más que con el largo del IPE per se. Por otra parte, el estudio de meta-análisis de Hess y col. (2005) indica la existencia de una correlación entre largo del IPE con la CC al momento del parto y con el balance de energía, pero no con la CC en el preparto. Entonces, los autores señalan que la CC al momento del parto resulta ser la única variable confiable a considerar para tener estimar los efectos del plano nutricional sobre el IPE.
Así entonces, los autores señalan que la CC al momento del parto resulta ser la única variable confiable a considerar para tener estimar los efectos del plano nutricional sobre el IPE.
Figura 2. Influencia del largo del intervalo posparto primer estro (IPE en días) sobre la tasa de preñez (Adaptado de Hess y col., 2005).
El crecimiento folicular después del parto ha sido asociado al consumo de energía y las reservas energéticas, ya que influyen en la concentración de substratos energéticos y hormonas metabólicas en la sangre (Wettemann y col., 1999). Así un bajo consumo de alimento implica baja en las reservas de grasa corporal que finalmente disminuyen las concentraciones de IGF-I, insulina y tiroxina (Lents y col., 2005). En consecuencia, el animal enfrenta un cuadro de balance de energía negativo. Cuando esto ocurre en el periodo posparto se observa un incremento en el número de folículos de tamaño medio y una disminución del diámetro del folículo dominante, mientras que un balance positivo de energía resulta en un aumento en el diámetro del folículo dominante y una reducción en el crecimiento de los folículos subordinados (Lucy y col., 1991). Por otra parte, un aumento en la cantidad de energía consumida después del parto resulta en un aumento del tamaño del folículo dominante en vacas primíparas y maduras (Ciccioli y col., 2003). Este mismo autor reporta que las vacas que presentan una CC moderada tienen folículos más grandes que vacas de CC delgada. Asimismo, el estudio realizado por Rhodes y col. (1995) señala que por cada 10 kg de pérdida de peso vivo el diámetro del folículo se reduce en 0,31(± 0,06) mm y el tamaño y persistencia del folículo dominante también se reduce.
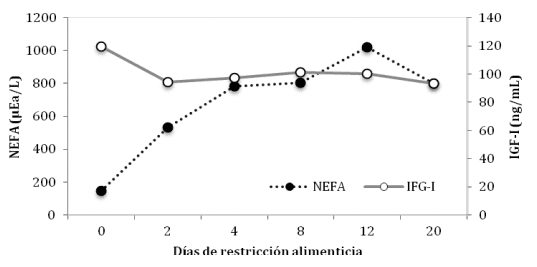
En una publicación reciente, Lents y col. (2011) diseñaron un experimento para evaluar el efecto de la ganancia diaria de peso y del consumo de alimento residual (Residual Feed Intake, RFI1 ) de vaquillas sobre la función lútea durante un periodo de subnutrición. En su experimento los autores no encontraron ninguna relación entre RFI y peso vivo, ganancia diaria de peso, consumo diario de alimento o espesor de la grasa dorsal. Tampoco encontraron relación entre RFI y concentración de IGF-I en el plasma. Sin embargo, todas las vaquillas perdieron peso (~47 kg) y redujeron el espesor de su grasa dorsal al finalizar la restricción alimenticia. Sobre un total de 67 vaquillas, 4 presentaron anestro y 16 tuvieron concentraciones de progesterona que fueron atípicas para un funcionamiento lúteo normal (~30%), durante el periodo de restricción alimenticia, el que correspondió a 21 días de una dieta que suministraba solamente el 40% de los requerimientos de energía. Por otra parte, las concentraciones de NEFA en plasma se incrementaron al segundo día de restricción alimenticia y permanecieron elevadas durante todo el periodo de subnutrición, mientras que la concentración de IGF-I presentó una respuesta completamente inversa (Figura 3).
Desde un punto de vista práctico la CC al momento del encaste es menos valiosa, como herramienta de manejo, que la CC al momento del parto, debido a que es muy difícil, sino imposible, el mejorar la CC una vez que el periodo de encaste ha comenzado. Además, existe una correlación positiva entre CC al momento de parto y la CC al momento del encaste (Hess y col., 2005). Finalmente, aun cuando los mecanismos precisos a través del cual los inputs nutricionales median o regulan la reproducción no han sido totalmente elucidados, existe consenso en que el eje reproductivo recibe señales nutricionales que lo afectan directa o indirectamente.
Figura 3. Concentración de IGF-I y NEFA en plasma de vaquillas durante un periodo de restricción alimenticia suministrando 40% de los requerimientos energéticos (Elaborado a partir de Lents y col, 2011).
Conclusión
La comprensión de los factores nutricionales asociados a IPE resultan claves a la hora de alcanzar la meta de parir un ternero(a) por vaca año. En este sentido, la utilización de tecnologías prácticas de bajo costo de implementación como lo es el monitoreo y registro de la CC y peso vivo al momento del parto se transforman en poderosas herramientas que en forma indirecta permiten evaluar el estado nutricional de los animales y mediante éste, su desempeño reproductivo. Si bien el balance energético de pre y postparto son los factores más relevantes afectando la duración del IPE, otros efectos nutricionales probablemente influyen en el eje hipotálamohipófisis-ovario que influyen en la reproducción. En Chile existen escasos antecedentes experimentales o de campo en el rubro carne que avalen estas investigaciones.
Literatura citada
Bossis, I., R.P. Wettemann, S.D. Welty, J.A. Vizcarra, L.J. Spicer, and M.G. Diskin. 1999. Nutritionally induced anovulation in beef heifers: Ovarian and endocrine function preceding cessation of ovulation. J. Anim Sci. 77:1536-1546.
Callejas S. 2001. Fisiología del ciclo estral bovino. En: Biotecnología de la Reproducción. Editado por G A. Palma. Ediciones Instituto Nacional de Tecnología Agropecuaria. Páginas 37-53.
Ciccioli, N.H., R.P. Wettemann, L.J. Spicer, C.A. Lents, F.J. White, and D.H. Keisler. 2003. Influence of body condition at calving and postpartum nutrition on endocrine function and reproductive performance of primiparous beef cows. J. Anim Sci. 81:3107-3120.
Correa-Orozco, A. y L.F. Uribe-Velásquez. 2010. La condición corporal como herramienta para pronosticar el potencial reproductivo en hembras bovinas de carne. Rev. Fac. Nal. Agr. Medellín 63(2): 5607-5619.
Dunn, T.G., and G.E. Moss. 1992. Effects of nutrient deficiencies and excesses on reproductive efficiency of livestock. J. Anim. Sci. 70:1580-1593.
DiCostanzo, A., J.E. Williams, and D.H. Keisler. 1999. Effects of short or long term infusions of acetate or propionate on luteinizing hormone, insulin, and metabolite concentrations in beef heifers. J. Anim. Sci. 77:3050-3056.
Hess, R.P. S. L. Lake, E. J. Scholljegerdes, T. R. Weston, V. Nayigihugu, J. D. C. Molle, and G. E. Moss. 2005. Nutritional controls of beef cow reproduction. J. Anim Sci. 83(E. Suppl.):E90–E106.
Lents, C.A., R.D. Randell, A.M. Stelzleni, L.C. Caldwell, and T.H. Welsh Jr. 2011. Function of corpus luteum in beef heifers is affected by acute submaintenance feeding but is not correlated with residual feed intake. J. Anim Sci. published on line July 15 2011. Disponible en: http://jas.fass.org/content/early/2011/09/07/ jas.2011-4097.
Lents, C.A., R.P. Wettemann, F.J. White, I. Rubio, N.H. Ciccioli, L.J. Spicer, D.H. Keisler, and M.E. Payton. 2005. Influence of nutrient intake and body fat on concentration of insulin-like growth factor-I, insulin, thyroxine, and leptin in plasma of gestating beef cows. J. Anim Sci. 83:586-596.
Lucy, M.C., C.R. Staples, F.M. Michel, and W.W. Thatcher. 1991. Energy balance and size and number of ovarian follicles detected by ultrasonography in early postpartum dairy cows. J. Dairy Sci. 74:473-482.
ODEPA. 2011. Boletín Carne bovina: tendencias de producción, precios y comercio exterior. 37 pág.
Richards, M.W., R.P. Wettemann, and H.M. Schoenemann. 1989. Nutritional anestrus in beef cows: Body weight change, body condition, luteinizing hormone and ovarian activity. J. Anim Sci. 67:1520-1526.
Rhodes, F.M., L.A. Fitzpatrick, K.W. Entwistle and G. De’ ath. 1995. Sequential changes in ovarian follicular dynamics in Bos indicus heifers before and after nutritional anestrous. J. Reprod. Fertil. 104:41-49.
Schillo, K.K. 1992. Effects of dietary energy on control of luteinizing hormone secretion in cattle an sheep. J. Anim Sci. 70:1271-1282.
Wettemann, R.P., and I. Bossis. 1999. Energy intake regulates ovarian function in beef cattle. Disponible en: http://www.asas.org/JAS/symposia/proceedings/0934.pdf. Visitado el 18 de Octubre 2011 (10 pág).
Wettemann, R.P., C.A. Lents, N.H. Ciccioli, F.J. White, and I. Rubio. 2005. Nutritional and suckling mediated anovulation in beef cows. J. Anim Sci. 81(E. Suppl. 2):E48-E59.
Temas relacionados:
Autores:

Universidad Austral de Chile
Recomendar
Comentar
Compartir

21 de mayo de 2019
Excelente enfoque desde el punto de vista de optimización de sistema productivo animal y el aprovechamiento de las oportunidades comerciales que se abren a nuestros países, en beneficio de nuestros productores.
Recomendar
Responder

¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.



Te puede interesar
