Cinética de la liberación ruminal de macrominerales en pasto kikuyo (Pennisetum clandestinum) cosechado a dos edades de rebrote
No hubo diferencia en el contenido ni en los parámetros de cinética de la liberación de estos minerales entre las muestras recolectadas a los 32 y 56 días (p>0.05). El K fue el mineral de mayor concentración (4.0%) seguido por el P (0.47%), el Ca (0.28%) y el Mg (0.26%). El Ca proveniente del rumen se fijó a las muestras del pasto durante los dos primeros tiempos de incubación con lo que la fracción soluble (a) para este mineral fue ligeramente negativa. Para los demás minerales se observó una fracción a que osciló entre el 70.4% para el Mg cosechado a los 58 días y 85.6% para el potasio cosechado a la misma edad. La fracción potencialmente liberable (b) fue alta para el Ca (91.3%) y baja para los otros tres minerales. El Ca presentó un retraso en el inicio de la fase rápida de la liberación que varió entre 5.02 y 7.70 horas. La liberación efectiva fue diferente entre los minerales siendo más baja en Ca (43.8%) y más alta en K (96.8%), e intermedia para Mg (84.6%) y P (90.4%). Los excesos en la concentración y liberación del K podría interferir en la utilización del Ca y el Mg.
KINETICS OF MACRO-MINERAL RELEASE IN THE RUMEN FRON KIKUYU GRASS (PENNISETUM CLANDESTINUM) HARVESTED TWO CUTTING INTERVALS
Abstract
To estimate the parameters of ruminal calcium (Ca), phosphorus (P), magnesium (Mg) and potassium (K) kinetics release from kikuyu grass (Pennisetum clandestinum) harvest at two cutting ages, an in situ test was developed utilizing four Holstein rumen cannulated females. Three samples of this grass were harvested at 32 and 58 of regrowth days from the same paddock and were analyzed for dry matter (DM), Ca, P, Mg and K. 28 sub-samples of 3.0 g approximately were taken from each sample. They were packed in nylon bags of 5 x 10 cm and they were incubated during seven times (0, 2, 6, 12, 24, 48, y 72 h) in the rumen of the four cannulated Holstein females (one bag by each time and by each sample). At end of incubation periods, the bags were washed with tap water and dried at 60oC during 48 h. The residues from the four bags of each incubation time and of each sample were mixed and were evaluated for DM, Ca, P, Mg and K. The percentage of rumen mineral release was estimated by difference between the initial and final mineral quantity in each nylon bag.
The cutting age did not affect the mineral content nor the ruminal mineral release kinetic parameters. K was the mineral with highest concentration (4.0%), followed by P (0.47%, Ca (0.28%) and Mg (0.26%). During the two first incubation times the rumen Ca was retained by grass samples and, in consequence, the soluble fraction (a) of this mineral was negative. For the other minerals a high a fraction was observed oscillating between 70.4% in Mg to 85.6% in K at harvests of 58 days of regrowth. The potentially releasable fraction (b) was high in Ca (91.3%) but low in the other minerals. Ca showed a delay in the beginning of the quick phase of release oscillating between 5.02 and 7.79 h. The effective ruminal release was different among all minerals being among lowest in Ca (43.8%) and highest in K (96.8%), and intermediate in Mg (84.6%) and in P (90.4%). The excess K content and effective ruminal release in kikuyu grass may affect the Ca and Mg utilization.
Key words: kikuyu grass, macrominerals, ruminal kinetics
La proteína microbiana puede aportar hasta dos tercios de los aminoácidos que son absorbidos en el intestino de los rumiantes (Verbic 2002). Es por ello que uno de los principales objetivos en los sistemas de alimentación de estos animales es optimizar la síntesis de proteína microbiana en el rumen (Vaughan et al 2002). Este proceso se ve afectada, entre otros factores, por la disponibilidad oportuna y suficiente de nutrientes y energía. Aunque ha sido más intensa la investigación sobre el efecto de la energía y la proteína fermentables en el rumen sobre este proceso (Verbic 2002), el papel que juegan los minerales no es menos importante (Samaniego 1996).
Se ha reconocido que la sincronía en la disponibilidad de nutrientes y energía es un componente importante en el diseño de programas de alimentación para rumiantes (Bach 2001; Vaughan et al 2002; Rémond et al 2002). Para ello, sin embargo, es necesario contar con información sobre los parámetros de cinética ruminal siendo la técnica in situ la más utilizada para estimar dichos parámetros (NRC 2001).
Dado que el pasto kikuyo (Pennisetum clandestinum) es el más utilizado en los sistemas de producción de leche bovina en el trópico alto en Colombia (Ministerio de Agricultura y Desarrollo Rural 1998), es importante estimar la concentración y los parámetros de cinética de la liberación ruminal de algunos macrominerales y su variación con la edad de corte con la finalidad de incorporar dicha información en la elaboración de programas de suplementación con estos minerales.
Es por ello que el objetivo de este trabajo fue estimar la concentración y los parámetros de cinética de la liberación ruminal del calcio (Ca), fósforo (P, magnesio (Mg) y potasio (K) en muestras de pasto kikuyo cosechadas a los 32 y 58 días de rebrote.
MATERIALES Y MÉTODOS
El trabajo de campo se llevó a cabo en el Centro de Producción Paysandú de la Universidad Nacional de Colombia, localizado en el corregimiento de Santa Elena (Medellín, Antioquia), a 2300 msnm, con una temperatura promedio de 16°C, perteneciente a una zona ecológica de bH - MB. En este Centro de Producción existe un programa de producción de leche, con animales de la raza Holstein y sus cruces con animales de la raza Blanco Orejinegro - BON, para lo cual dispone de aproximadamente 70 hectáreas distribuidas en 94 potreros, la mayor parte de los cuales se encuentran sembrados con pasto kikuyo (Pennisetum clandestinum) en diferentes estados de rebrote. Aproximadamente la mitad de los potreros se encuentran asignados a las vacas en producción, los cuales son manejados en un sistema de pastoreo por franjas y son fertilizados con 25-15-0-2-3 a razón de 200 kg/ha luego de cada pastoreo.
Tres muestras de este pasto provenientes de un mismo potrero fueron recolectadas a los 32 y a los 58 días (aproximadamente 2.0 kg de materia verde/muestra). Estas muestras fueron secadas a 65oC por 48 horas, a las cuales se les determinó el contenido de cenizas totales (Cen), Ca, P, Mg y K. Así mismo, se molieron en criba de 1.5 mm, se empacaron en bolsas de nilón de 5 x 10 cm (aproximadamente 3.0 gr/bolsa) y se incubaron durante 0, 2, 6, 12, 24, 48 y 72 horas en el rumen de cuatro vacas Holstein canuladas, utilizando una bolsa para cada tiempo en cada animal. Las bolsas se sujetaron a mosquetones metálicos, los cuales, a su vez, se sujetaron a una cadena metálica de un metro de longitud que se introdujo en la parte ventral del rumen y se sujetó a la parte externa de la tapa de la cánula ruminal, mediante un cordel de cáñamo. La bolsas se incubaron en el rumen comenzando con el tiempo más largo (72 horas) prosiguiendo hasta las 0 horas (20 minutos), de tal manera que al final de la incubación, todas las bolsas se extrajeron al mismo tiempo y se lavaron simultáneamente con agua de grifo hasta que el agua salió limpia (Nocek 1988).
Las bolsas con los residuos se secaron a 65°C durante 48 horas, se colocaron al ambiente hasta que su temperatura se estabilizó y se pesaron. En el residuo que quedó en cada bolsa incubada se determinó el contenido de MS, Ca, P, Mg y K, cuyos datos se utilizaron para estimar los parámetros de la cinética de liberación ruminal a partir de la ecuación de Mitcherlish I (Kiviste et al 2002), para lo cual se utilizó el programa RUMENAL (Correa 2004). Se utilizó el PROC TTEST del programa estadístico SAS (1998) para establecer las diferencias entre la edad de corte, para los parámetros estimados. Finalmente, se calculó la liberación efectiva de los minerales mediante el procedimiento propuesto por Ørskov y McDonald (1979) asumiendo una kp de 0.05/h y recurriendo al PROC GLM del programa estadístico SAS (1998) para establecer las diferencias entre los minerales para este parámetro.
RESULTADOS Y DISCUSIÓN
Contenido de Cen, Ca, P, Mg y K
En la tabla 1 se presentan los resultados del contenido de Cen, Ca, P, Mg y K de las muestras del pasto kikuyo cosechadas a los 32 y 58 días de rebrote.
Tabla 1. Contenido de Ceniza, Ca, P, Mg y K del pasto kikuyo cosechado a 32 (kik32) y a 58 días de rebrote (kik58).
Fracción | kik32 | kik58 | p |
Ceniza | 13.1 | 12.8 | 0.54 |
Ca | 0.29 | 0.26 | 0.18 |
P | 0.50 | 0.44 | 0.13 |
K | 4.12 | 3.96 | 0.30 |
Mg | 0.25 | 0.26 | 0.15 |
Como se puede apreciar, la edad de corte no afectó ninguno de los parámetros evaluados (p>0.05). En general, el contenido de Cen fue más alto que el reportado por Soto et al (2005) pero similar al hallado por Gaitán y Pabón (2003). El contenido de Ca y Mg fueron menores a los valores reportados por Laredo (1988) y por Dugmore (1998) para esta gramínea. Por el contrario, el contenido de P y de K fueron más altos que los reportados por estos autores. El incremento en el contenido de estos minerales puede ser debido a la aplicación de fertilizantes que presentan altas concentraciones de estos minerales, pero menores concentraciones de minerales como el Ca y el Mg. Altos contenidos de P pueden afectar la absorción de Ca y Mg (Van Soest 1994; Underwood y Suttle 1999) requiriendo hacer las correcciones correspondientes a través de la suplementación mineral. Dugmore (1998) señala, además, que en la medida en que la relación K/(Ca + Mg) se incremente por encima de 2.2 (sobre una base iónica equivalente), en esa medida se incrementa la posibilidad de presencia de hipocalcemia. En el presente trabajo y teniendo en cuenta la ecuación propuesta por este autor (%K x 25.6)/(%Ca x 49.9 + %Mg x 82.3), se estimó que dicha relación asciende a 3.0 para el pasto recolectado a los 32 días y a 2.94 para el recolectado a los 58 días.
CURVAS DE LIBERACIÓN RUMIAL DE MINERALES
Calcio
En la figura 1 se muestran las curvas de cinética ruminal del Ca en el pasto kikuyo cosechado a los 32 (figura 1a) y a los 58 días (figura 1b) de rebrote.
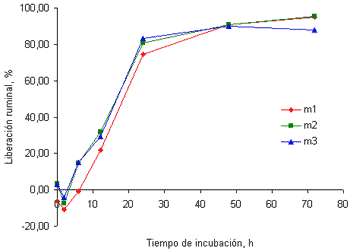
Figura 1a. Curvas de liberación del Ca de las tres muestras de pasto kikuyo cosechado a los 32 dias de rebrote.
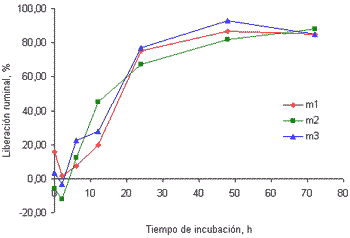
Figura 1b. Curvas de liberación del Ca de las tres muestras de pasto kikuyo cosechado a los 58 días de rebrote.
Como se puede apreciar, en los primeros tiempos de incubación se presentaron datos de liberación negativa que indican incrementos en la concentración de este mineral en los residuos recuperados, esto es, que parte del Ca disuelto en el rumen fue atrapado por el material incubado en las bolsas de nilón. Este comportamiento fue diferente al observado en los otros minerales evaluados en este trabajo. Este fenómeno parece estar relacionado con el secuestro de este mineral por las paredes celulares o por el ácido oxálico (Marais 1998). Emanuele et al (1991) reportaron que el Ca puede ser secuestrado por las paredes celulares dependiendo del pH del medio y del nivel de degradación de las fracciones fibrosas de los forrajes. Estos autores evaluaron la liberación del Ca en el intestino en muestras de dos leguminosas y cuatro gramíneas, luego de la incubación por 24 horas en el rumen y una hora en una solución de HCl/Pepsina, encontrando que la concentración del Ca en los residuos tras la incubación intestinal fue mayor indicando el secuestro de este mineral por la fracción fibrosa de los forrajes. Estos resultados indican, además, que la capacidad de retención del Ca en mayor en las gramíneas que en las leguminosas, esto es, que es mayor en los forrajes con mayor contenido de fibra en detergente neutro.
Marais (1998; 2001), por su parte, señala que el pasto kikuyo puede acumular cantidades importantes de ácido oxálico y que este compuesto tiene una alta capacidad de hacer insolubles cationes divalentes como el Ca, lo cual podría ser parcialmente responsable de la retención inicial del Ca en el rumen. Pasadas las primeras dos horas de incubación, sin embargo, se da inicio al proceso de liberación del Ca hasta alcanzar valores cercanos al 93% a las 72 horas de incubación, en las muestras recolectadas a los 32 días de rebrote. Este comportamiento sería explicable en los términos que sugieren Emanuele et al (1991), esto es, que en la medida en que se incrementa la degradación de la fibra, en esa medida se incrementa la liberación del Ca. Por su parte Allison et al (1977) señalan que los animales que consumen forrajes que acumulan cantidades significativas de ácido oxálico, como el pasto kikuyo, presentan un incremento en la población de Oxalobacter formigenes, una bacteria anaerobia obligada, que degrada este compuesto a formato y dióxido de carbono. Debido a que los cristales de oxalato de Ca se asocian con la porción fibrosa de las plantas, al incrementarse la degradación de la fibra se incrementa la degradación de este compuesto y, por ende, la liberación del Ca en el rumen (Franceschi y Horner 1980).
La liberación del Ca en el rumen de fuentes con bajo contenido de fibra presenta un comportamiento muy diferente al hallado en este trabajo. Brown et al(2004) reportaron las curvas de liberación ruminal de varios minerales, incluido el Ca, en muestras de maíz y semilla de algodón, en las que no se aprecia la retención de este mineral por los residuos, tras la incubación por tres horas, como sí se encontró en este trabajo, luego de dos horas de incubación en el rumen. Estos resultados reforzarían mucho más la idea de que en la medida en que se incrementa el contenido de FDN, mayor es la retención del Ca y menor su disponibilidad, tanto a nivel ruminal como a nivel posruminal.
Fósforo
Al contrario de lo encontrado con el Ca, las curvas de liberación del P en el rumen, muestran que este mineral es de muy alta disponibilidad en el rumen. La liberación de este mineral luego de 20 minutos de incubación en el rumen (tiempo 0) fue superior al 80% alcanzando valores cercanos al 98% a las 72 horas de incubación (figuras 2a y 2b). Esto puede ser debido a que el P se distribuye uniformemente entre las hojas y los tallos de las gramíneas (Underwood y Suttle 1999) y hay baja formación de ácido fítico, contrario a lo que sucede con cereales (Nelson et al 1976). Aunque el P fítico es de baja disponibilidad para los no rumiantes (NRC 1998) la alta actividad fitasa en el rumen hace disponible este mineral (Godoy y Meschy 2001; Nelson et al 1976) explicando en cierta medida, la liberación tan alta del P hallada en este trabajo.
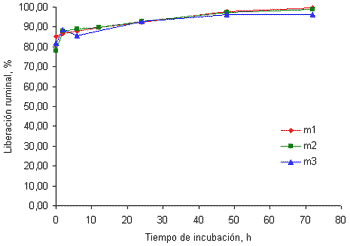
Figura 2a. Curvas de liberación del P de las tres muestras de pasto kikuyo cosechado a los 32 días de rebrote
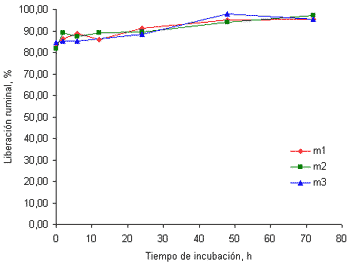
Figura 2b. Curvas de liberación del P de las tres muestras de pasto kikuyo cosechado a los 58 días de rebrote
La liberación ruminal del P en el tiempo cero en alimentos concentrados es más baja que los valores hallados en este trabajo. Así, Brown et al(2004) encontraron que la liberación inicial del P fue de 55.8% en harina maíz y de 11.8% en torta de algodón. La liberación de este mineral en los cereales mencionados, sin embargo, se incrementó hasta alcanzar valores superiores al 90% a las 24 horas de incubación. Estos valores son similares a los hallados en el presente trabajo.
Magnesio
Este mineral presentó un comportamiento similar al del P, aunque con una liberación inicial menor (cercana al 70%) (figuras 3a y 3b).
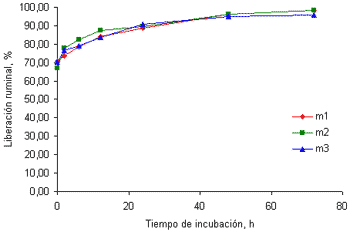
Figura 3a. Curvas de liberación del Mg de las tres muestras de pasto kikuyo cosechado a los 32 días de rebrote.
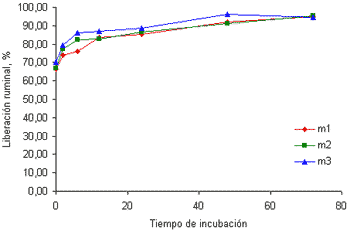
Figura 3b. Curvas de liberación del Mg de las tres muestras de pasto kikuyo cosechado a los 58 días de rebrote.
Esto podría estar relacionada con el efecto del pH ruminal sobre la solubilidad de este mineral. Según la NRC (2001) en la medida en que el pH ruminal se incremente por encima de 6.5, se reduce la solubilidad de este mineral y es precisamente un pH de este tenor el que podría esperarse en animales consumiendo forrajes con altos contenidos de PC y K como los hallados en este trabajo. El contenido de PC en las muestras recolectadas a los 32 días fue de 21.5% y fue de 16.6% en las muestras recolectadas a los 58 días. Este tipo de forrajes están acompañados de bajos contenidos de carbohidratos no estructurales (Soto et al 2005) que no permiten que el amonio formado durante la fermentación de las proteínas se fije a la proteína microbiana, generando una alta concentración de amonio. Este ión junto con el K, incrementan el pH ruminal y reducen, en consecuencia la solubilidad y la liberación inicial del Mg en el rumen.
Emanuele et al (1991) encontraron que la liberación del Mg en cuatro gramíneas y dos leguminosas, luego de 24 horas de incubación en el rumen, osciló entre 87.8 y 97.7%, valores cercanos a los hallados en este trabajo. Por el contrario, la liberación del Mg en harina de maíz y torta de algodón es mucho menor que la encontrada en forrajes según los datos hallados por Brown et al(2004).
Potasio
El K no solo es el mineral en mayor concentración (tabla 1) si no que, además, es el que presenta la mayor disponibilidad a nivel ruminal.
La liberación del K en el tiempo cero osciló entre 83 y 85% para las muestras recolectadas a los 32 (figura 4a) y 58 días (figura 4b), respectivamente.
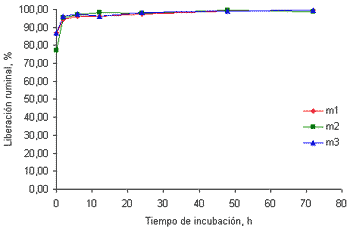
Figura 4a. Curvas de liberación del K de las tres muestras de pasto kikuyo cosechado a los 32 días de rebrote.
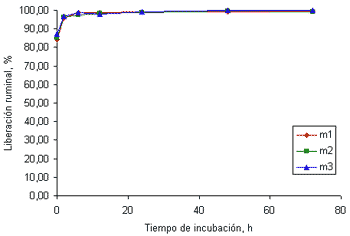
Figura 4b. Curvas de liberación del K de las tres muestras de pasto kikuyo cosechado a los 58 días de rebrote.
las 24 horas de incubación la liberación de este mineral alcanzó valores superiores al 99%. Estos resultados concuerdan con los valores reportados por Emanuele et al (1991) quienes encontraron que la liberación de este mineral en el rumen en cuatro gramíneas y dos leguminosas luego de 24 horas de incubación osciló entre 99.7 y el 100%. Los resultados reportados por Brown et al(2004) indican que la liberación inicial de este mineral es ligeramente menor en harina de maíz (87.3%) y en torta de algodón (85.1%) que en gramíneas. Aún así, la liberación de este mineral en el rumen alcanza valores cercanos al 100% a las 24 horas de incubación.
PARÁMETROS DE CINÉTIVA DE LIBERACIÓN RUMINAL DE MINERALES
Los parámetros de cinética ruminal permiten hacer inferencias sobre algunas de las características que presentan las fracciones nutricionales que se degradan o liberan en el rumen. El valor que se obtenga de cada parámetro va a depender del modelo matemático utilizado (Naranjo et al 2005). Estos autores evaluaron la capacidad de ajuste de cuatro modelos matemáticos, tres de los cuales han sido ampliamente utilizados para estimar los parámetro de cinética ruminal. El modelo de Mitscherlich I modificado (Kiviste et al 2002) no obstante no haber sido utilizado previamente para estos fines, demostró ser el que mejor se ajusta los datos de cinética ruminal independientemente de la fracción nutricional y de las características de los datos analizados, permitiendo describir de manera más apropiada los procesos biológicos involucrados en la degradación ruminal, . Es por ello que en este trabajo se utilizó este modelo para estimar los parámetros de cinética de la liberación ruminal de los minerales.
La tabla 2 muestra los parámetros de la cinética de la liberación del Ca en las muestras del pasto kikuyo cosechado a los 32 y a los 58 días. No hubo diferencia debidas a la edad de corte (p>0.05) para ninguno de los parámetros. Como se puede apreciar, la fracción a presentó valores negativos, debido fundamentalmente a la retención que sufrió este mineral durante las dos primeras horas de incubación en el rumen, como fue discutido previamente. La fracción b fue bastante alta, indicando que una alta proporción de este mineral es potencialmente degradado en el rumen.
Tabla 2. Parámetros de la cinética de liberación ruminal del Ca en pasto kikuyo cosechado a los 32 y 58 días de rebrote.
Parámetros | 32 d | 58 d | p |
a | -1.87 | -1.25 | 0.91 |
b | 94.5 | 88.2 | 0.30 |
kd | 0.14 | 0.14 | 0.93 |
i | 5.02 | 7.69 | 0.64 |
scep* | 111 | 133 | 0.81 |
La relación entre la aceleración y la velocidad de degradación (kd) (Correa 2005) es mayor que los valores reportados por Naranjo et al (2005) para la materia seca, la proteína cruda y la fibra en detergente neutro en algunas forrajeras tropicales y que los valores reportados por Brown et al(2004) para harina de maíz y torta de algodón. El punto de inflexión (i) es el tiempo en el que comienza el crecimiento exponencial de la curva de degradación (Solano y Vargas 1997) y equivale al tiempo de retraso o tiempo lag utilizado en otros modelos (McDonald 1981; Mertens y Ely 1982). Para el caso del Ca, este parámetro varió entre 5.02 y 7.70 horas indicando que la liberación rápida del Ca se logra después de transcurrido este tiempo y correspondería a la degradación rápida de la fibra.
Los parámetros de cinética de la liberación ruminal del fósforo tampoco fueron diferentes entre las muestras de kikuyo recolectadas a los 32 o a los 58 días (tabla 3).
Tabla 3. Parámetros de la cinética de liberación ruminal del P en pasto kikuyo cosechado a los 32 y 58 días de rebrote.
32 d | 58 d | p | |
a | 83.06 | 84.59 | 0.31 |
b | 16.20 | 14.69 | 0.24 |
kd | 0.05 | 0.02 | 0.12 |
i | 1.09 | 1.00 | 0.37 |
scep* | 13.39 | 20.04 | 0.65 |
A diferencia del Ca, la fracción soluble del P (fracción a) fue muy alta, reflejando la liberación de este mineral en el tiempo cero. La fracción b por el contrario, fue menor que la hallada para el Ca, lo mismo que la kd. Esto indica una menor facilidad en la liberación de esta fracción del P en comparación a la encontrada para el Ca. Es probable que se trate de P ligado a fracciones orgánica de menor degradación, tales como la fibra en detergente neutro o la proteína ligada a la fibra en detergente neutro. El valor de 1.0 en el punto de inflexión indica que no existe un tiempo de retraso para el inicio de la fase rápida de la liberación de este mineral.
El Mg presentó una fracción a mayor que la del Ca, pero un poco menor que la del P. La fracción b y la kd, por el contrario, fueron mayores que la del P (tabla 4).
Tabla 4. Parámetros de la cinética de liberación ruminal del Mg en pasto kikuyo cosechado a los 32 y 58 días de rebrote.
32 d | 58 d | p | |
a | 71.06 | 70.41 | 0.54 |
b | 26.38 | 22.89 | 0.12 |
kd | 0.07 | 0.10 | 0.34 |
i | 1.00 | 1.00 | - |
scep* | 17.70 | 41.21 | 0.20 |
Los valores encontrados para la fracción a y la b del K fueron similares a los valores hallados para el P. La kd del K fue, sin embargo, mucho más alta en el K que la estimada en el P y que la de los otros dos minerales (tabla 5). Esto indica que la liberación total del K es mayor y más rápida que la que presentan los demás minerales, poniendo en evidencia la posible interacción negativa entre la cantidad de K disponible en el rumen y la utilización de los demás minerales (NRC 2001).
Tabla 5. Parámetros de la cinética de liberación ruminal del K en pasto kikuyo cosechado a los 32 y 58 días de rebrote.
32 d | 58 d | p | |
a | 83.73 | 85.60 | 0.60 |
b | 14.39 | 13.38 | 0.75 |
kd | 0.77 | 0.81 | 0.79 |
i | 1.00 | 1.00 | - |
Sep* | 6.74 | 1.68 | 0.05 |
LIBERACIÓN RUMINAL EFECTIVA DE MINERALES
La liberación efectiva de los minerales no fue diferente entre las muestras cosechadas a los 32 y a los 58 días, pero sí fue diferente entre minerales (p<0.0001) siendo, en su orden, K>P>Mg>Ca (tabla 6).
Tabla 6. Liberación efectiva de Ca, P, Mg y K en pasto kikuyo cosechado a los 32 y 58 días de rebrote.
kik32 | kik58 | Media | p | |
Ca | 43.97 | 43.39 | 43.84d | 0.92 |
P | 90.70 | 90.02 | 90.36b | 0.41 |
Mg | 85.16 | 83.97 | 84.57c | 0.40 |
K | 96.33 | 97.27 | 96.80a | 0.04 |
p | 0.0001 |
Este parámetro refuerza aún más la idea de que los excesos de K pueden estar afectando la disponibilidad de Mg, pero sobre todo de Ca en los sistemas de producción bovina basados en pastos como el kikuyo. Estos resultados demuestran, además, la necesidad de seguir explorando la cinética de la liberación de minerales en el rumen bajo metodologías como la utilizada en este experimento.
CONCLUCIONES
Los parámetros de la cinética de la liberación ruminal de minerales en pasto kikuyo no fue diferentes entre las muestras tomadas a los 32 y a los 58 días de rebrote.
Se demostró, sin embargo, que la cinética de la liberación ruminal es diferente entre los minerales evaluados siendo el Ca el que presentó la menor liberación efectiva y el K la mayor liberación efectiva.
- Este comportamiento tiene importantes implicaciones en cuanto a la disponibilidad de los minerales en el rumen.
AGRADECIMIEMTOS
A la Dirección de Investigaciones de la Universidad Nacional de Colombia, Sede Medellín (DIME) por la financiación de este trabajo (proyecto DIME 030803758) y a los auxiliares de investigación Ferley Monsalve y Martha Ruíz por la excelente labor realizada en el montaje y la ejecución del trabajo de campo.
REFERENCIAS
Allison M J, Littledike E T and James L F 1977 Changes in ruminal oxalate degradation rates associated with adaptation of oxalate ingestion; Journal of Animal Science (45): 173.
Bach A 2001 Sincronización de la disponibilidad de nitrógeno y carbohidratos en el rumen; Frisona Española (121, Enero/Febrero): 84.
Brown M S, Drager C D, Cochran E M and Lauterbach E A 2004 Ruminal mineral release from corn and cotton seed meal. TX and Texas Agricultural Experiment Station, West Texas A and M University, Division of Agriculture. 4 p.
Correa H J 2004 Rumenal: procedimiento para estimar los parámetros de cinética ruminal mediante la función Solver de Microsoft Excel®. Revista Colombiana de Ciencias Pecuarias. Volumen 17 (3): 250.
Correa H J 2005 Origin and comprehensive study of Thünen´s model to analyze data from in situ rumen degradability technique. Revista Colombiana de Ciencias Pecuarias, Volumen 18 (1): 80.
Dugmore T J 1998 Energy and mineral content of kikuyu; In: Bartholomew P E (Editor) Proceedings of a Kikuyu Technology Day, held at Cedara on 25 November, KwaZulu-Natal Department of Agriculture 16.
Emanuele S M, Staples C R and Wilcox C J 1991 Extent and site of mineral release from six forage species incubated in mobile dacron bags; Journal of Animal Science (69): 801
Franceschi V R and Horner H T 1980 Calcium oxalate crystals in plants; Botanical Review (46): 361.
Gaitán S y Pabón J D 2003 Aplicación del modelo NRC 2001 en la caracterización energética y proteica de los pastos kikuyo (Pennisetum clandestinum, Hochst), ryegrass (Lolium perenne) y falsa poa (Holcus lanatus,) en un hato lechero del oriente antioqueño; Trabajo de Grado de Zootecnia, Departamento de Producción Animal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. 55 p.
Godoy S and Meschy F 2001 Utilization of phytate phosphorus by rumen bacteria in a semi-continuous culture system (Rusitec) in lactating goats fed on different forage to concentrate ratios; Reproduction and Nutrition Development. (41): 259.
Kiviste A, Álvarez J G, Rojo A y RuízA D 2002 Funciones de crecimiento de aplicación en el ámbito forestal; INIA. Madrid, España. 190 p.
Laredo M A 1988 Tabla de contenido nutricional en pastos y forrajes de Colombia; Medellín: ICA – Colanta. 40 pg.
Marais J P 1998 Anti quality factors; In: Bartholomew P E (Editor) Proceedings of a Kikuyu Technology Day, held at Cedara on 25 November, KwaZulu-Natal Department of Agriculture: 21.
Marais J P 2001 Factors affecting the nutritive value a kikuyu grass (Pennisetum clandestinum) - a review; Tropical Grassland (35): 65.
McDonald I 1981 A revised model for the estimation of protein degradability in the rumen; Journal of Agricultureal Science (Cambridge) (96): 251.
Mertens D R y Ely L O 1982 Relationship of rate and extent of digestion to forage utilization: A dynamic model evaluation; Journal of Animal Science (54): 895.
Ministerio de Agricultura y Desarrollo Rural 1998 Acuerdo Nacional de Competitividad de la Cadena Láctea; Santafé de Bogotá, D. E. 123 pg.
Naranjo J F, Cuartas C A y Correa H J 2005 Comparación de cuatro modelos matemáticos para la caracterización de la cinética de degradación ruminal de algunos recursos forrajeros. Livestock Research for Rural Development. Volume 17, Article #98 Retrieved September 1, 2005 http://www.cipav.org.co/lrrd/lrrd17/9/ nara17098.htm
N R C (National Research Council 1998) Nutrient Requirements of Swine; 10th revised edition, National Academy Press, Washington, D. C.
N R C (National Research Council 2001) The nutrient requirement of dairy cattle; Seventh edition. National Academy Press, Washington, D. C.
Nelson T S, Daniels L B, Hall J R and Shields L G 1976 Hydrolysis of natural phytate phosphorus in the digestive tract of calves; Journal of Animal Science (42):1509.
Nocek J E 1988 In situ and other methods to estimate ruminal protein and energy digestibility: A review; Journal of Dairy Science (71): 2051.
Ørskov E R and McDonald I 1979 The estimate of protein degradability in the rumen from incubation measurements weighed according to rate of passage; Journal of Agricultural Science (Cambridge) (92): 499.
Rémond D, Nozière P and Poncet C 2002 Effect of time of starch supply to the rumen on the dynamics of urea and ammonia net flux across the rumen wall of sheep; Animal Research (51): 3.
Samaniego M A 1996 Comparison of methods for the measurement of digesta flow and microbial protein supply in sheep; MSc. Thesis, University of Aberdeen. 57 pg.
SAS Inst 1998 SAS User's Guide: Statistics (Version 8); Cary NC: the Institute.
Solano C y Vargas L 1997 El crecimiento de novillas de reemplazo en fincas lecheras de Costa Rica. 1. Tipificación del crecimiento de novillas Holstein y Jersey; Archivos Latinoamericanos de Producción Animal (5): 21.
Soto C, Valencia A, Galvis R D y Correa H J 2005 Efecto de la edad de corte y del nivel de fertilización nitrogenada sobre el valor energético y proteico del pasto kikuyo (Pennisetum clandestinum). Revista Colombiana de Ciencias Pecuarias. Volumen 18 (1): 17.
Underwood E J and Suttle N F 1999 The mineral nutrition of livestock; 3rd Edition. CABI publishing, Wallingford, UK. 600 p.
Van Soest P J 1994 Nutritional ecology of the ruminant; Cornell University Press. 476 p.
Vaughan J M, Bertrand J A, Jenkins T C and Pinkerton B W 2002 Effects of Feeding Time on Nitrogen Capture by Lactating Dairy Cows Grazing Rye Pasture; Journal of Dairy Science (85):1267.
Verbic J 2002 Factors affecting microbial protein synthesis in the rumen with emphasis on diets containing forages; In: Viehwirtschaftliche Fachtagung, BAL Gumpenstein. 6 p.






