Situación Mundial de la Influenza Aviar y sus implicaciones en América Latina
Publicado: 13 de octubre de 2010
Por: Miguel Márquez, United States Soybean Export Council American Soybean Association-IM. Consultor Internacional. Guadalajara, México. Julio 2010
Introducción
La Ortomixovirosis o Influenza Aviar de Alta Patogenicidad en aves domésticas, ha sido reportada con una frecuencia creciente a lo largo de los últimos 55 años en las industrias avícolas de diferentes países y en diversos continentes, proceso preocupante y difícil de explicar por veterinarios clínicos, avipatólogos, virólogos, inmunólogos y epidemiólogos. Asimismo, la constante circulación viral, con la aparición y desaparición del agente de la Influenza Aviar de Baja Patogenicidad, tanto en parvadas de pollo (Gallus gallus domesticus), como de pavos (Meleagris gallopavo), denota la gran actividad y plasticidad genética de estos ortomixovirus.
Por otro lado, el concepto de fronteras nacionales entre los países parece ser ya no más válido, debido al nuevo comportamiento viral el cual debe ser contemplado ahora como "global" desde un punto de vista geográfico, dada su intensa movilidad intra e intercontinental.
La Ortomixovirosis o Influenza Aviar de Alta Patogenicidad en aves domésticas, ha sido reportada con una frecuencia creciente a lo largo de los últimos 55 años en las industrias avícolas de diferentes países y en diversos continentes, proceso preocupante y difícil de explicar por veterinarios clínicos, avipatólogos, virólogos, inmunólogos y epidemiólogos. Asimismo, la constante circulación viral, con la aparición y desaparición del agente de la Influenza Aviar de Baja Patogenicidad, tanto en parvadas de pollo (Gallus gallus domesticus), como de pavos (Meleagris gallopavo), denota la gran actividad y plasticidad genética de estos ortomixovirus.
Por otro lado, el concepto de fronteras nacionales entre los países parece ser ya no más válido, debido al nuevo comportamiento viral el cual debe ser contemplado ahora como "global" desde un punto de vista geográfico, dada su intensa movilidad intra e intercontinental.
El creciente número de brotes reportados debidamente documentados, provocados por virus de Influenza Aviar de Alta Patogenicidad (IAAP) desde mediados del siglo pasado, demuestra una intensa activación de esta enfermedad, la cual esta causando de más en más, un mayor número de brotes, los cuales toman carácter epizoótico y algunos de entre estos episodios adquieren proporciones de panzootias, provocando como es el caso de la Influenza Aviar por virus H5N1 en el Sureste Asiático el cual ha producido enormes pérdidas económicas por la mortalidad misma y por los severas medidas de erradicación con sacrificio de los animales, además de la terrible y cuantiosa penuria pecuniaria, resultante de las medidas cuarentenarias y de bioseguridad, tales como, la restricción de movilización, control de la comercialización y las barreras sanitarias intraestatales o bien internacionales entre países, que terminan arruinando la economía avícola y pecuaria de las naciones afectadas.
Desde 1955, han sido científicamente documentados ante la Office International des Epizooties (OIE) de París, actualmente conocida como Organización Mundial de Salud Animal (OMSA), numerosos brotes de Influenza Aviar (IA) causados por virus de Alta Patogenicidad, es decir, que durante ultimas seis décadas se han presentado, un promedio de 1.7 brotes por año en diferentes continentes y lo más preocupante es que las últimos veinte años, estas epizootias han ocurrido a lo largo de la década de los años 90 del siglo pasado y la primera década del presente siglo, esto aunado a los cientos de virus de Baja Patogenicidad que han sido aislados durante los últimos 60 años.
En la mayoría de los casos, y cuando el diagnóstico ha sido confiable y precoz porque han existido los medios técnicos y sobretodo, económicos, la IA ha sido controlada por medio de la erradicación, gracias al sacrificio total de las aves afectadas y de las no afectadas, en estrecha vecindad. Sin embargo, en algunos de estos países, particularmente donde históricamente se optó por la vacunación para el control y la prevención de esta enfermedad altamente infecto-contagiosa, ella ha tomado desgraciadamente un carácter crónico-enzoótico.
Todo esto indica, entonces, que los virus causantes de la Influenza Aviar de Alta y de Baja Patogenicidad, en los próximos años incrementarán su presencia y acelerarán su virulencia en la avicultura industrial de países desarrollados y en naciones en desarrollo, por igual, debido al intenso intercambio comercial, resultado y consecuencia directa de numerosos tratados de libre comercio, entre bloques de países no forzosamente vecinos e igualmente a causa de la globalización económica y cultural del mundo y a la intensa movilización internacional de productos avícolas, gracias a los modernos y rápidos medios de transportación, así como, la diaria trashumancia de millones de seres humanos en perpetuo, multidinario y acelerado movimiento aéreo, sin descartar por supuesto, la posibilidad y el riesgo que representa, la migración estacional de aves acuáticas y terrestres.
Desde 1955, han sido científicamente documentados ante la Office International des Epizooties (OIE) de París, actualmente conocida como Organización Mundial de Salud Animal (OMSA), numerosos brotes de Influenza Aviar (IA) causados por virus de Alta Patogenicidad, es decir, que durante ultimas seis décadas se han presentado, un promedio de 1.7 brotes por año en diferentes continentes y lo más preocupante es que las últimos veinte años, estas epizootias han ocurrido a lo largo de la década de los años 90 del siglo pasado y la primera década del presente siglo, esto aunado a los cientos de virus de Baja Patogenicidad que han sido aislados durante los últimos 60 años.
En la mayoría de los casos, y cuando el diagnóstico ha sido confiable y precoz porque han existido los medios técnicos y sobretodo, económicos, la IA ha sido controlada por medio de la erradicación, gracias al sacrificio total de las aves afectadas y de las no afectadas, en estrecha vecindad. Sin embargo, en algunos de estos países, particularmente donde históricamente se optó por la vacunación para el control y la prevención de esta enfermedad altamente infecto-contagiosa, ella ha tomado desgraciadamente un carácter crónico-enzoótico.
Todo esto indica, entonces, que los virus causantes de la Influenza Aviar de Alta y de Baja Patogenicidad, en los próximos años incrementarán su presencia y acelerarán su virulencia en la avicultura industrial de países desarrollados y en naciones en desarrollo, por igual, debido al intenso intercambio comercial, resultado y consecuencia directa de numerosos tratados de libre comercio, entre bloques de países no forzosamente vecinos e igualmente a causa de la globalización económica y cultural del mundo y a la intensa movilización internacional de productos avícolas, gracias a los modernos y rápidos medios de transportación, así como, la diaria trashumancia de millones de seres humanos en perpetuo, multidinario y acelerado movimiento aéreo, sin descartar por supuesto, la posibilidad y el riesgo que representa, la migración estacional de aves acuáticas y terrestres.
Etiología
Los virus responsables de la Influenza Aviar son capaces de provocar severas infecciones agudas en el caso de virus de alta patogeninicidad y crónicas, sobre todo, en el caso de virus de baja patogenicidad. Clínicamente cursan con cuadros caracterizados por neumo-entero-encefalitis. Los agentes de la Influenza Aviar forman parte de la gran familia Orthomixoviridae del Tipo A. Son partículas víricas envueltas por una capa bilipídica, que miden aproximadamente 100 nm, portan ácido ribonucleico (ARN) y su genoma esta dividido de ocho segmentos en sentido inverso.
Además, poseen unas remarcables proteínas de superficie, 16 hemogalutininas y 9 neuroaminidadas, lo que les confiere una enorme potencial de recombinacion de hasta 144 diferentes subtipos. De entre ellos, los subtipos más importantes en medicina aviar son los subtipos H5 y H7, sin embargo, el los virus H9 ostentan un potencial infectológico que no debemos perder de vista en el futuro cercano.
Además, poseen unas remarcables proteínas de superficie, 16 hemogalutininas y 9 neuroaminidadas, lo que les confiere una enorme potencial de recombinacion de hasta 144 diferentes subtipos. De entre ellos, los subtipos más importantes en medicina aviar son los subtipos H5 y H7, sin embargo, el los virus H9 ostentan un potencial infectológico que no debemos perder de vista en el futuro cercano.
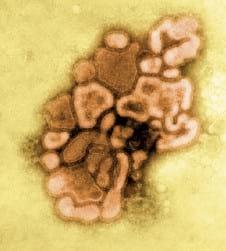
Virus de Influenza Aviar,
Egipto, 2010
Ahora bien, los subtipos H5 y H7 se pueden clasificar en virus de Alta y de Baja Patogenicidad.
La Organización Mundial para la Salud Animal (AMSA), mejor conocida como Oficina Internanacional de Epizootias (OIE), con sede en Paris, Francia, los define de la manera siguiente: "Para propósitos del Comercio Internacional los virus H5 y H7 de Influenza Aviar Notificable (IAN), son aquellos que son capaces de provocar la enfermedad en aves corral con IPIV >1.2 en pollos 6 semanas de edad o causar mortalidad >75% en pollos 4-8 semanas edad inoculados vía intravenosa".
La Organización Mundial para la Salud Animal (AMSA), mejor conocida como Oficina Internanacional de Epizootias (OIE), con sede en Paris, Francia, los define de la manera siguiente: "Para propósitos del Comercio Internacional los virus H5 y H7 de Influenza Aviar Notificable (IAN), son aquellos que son capaces de provocar la enfermedad en aves corral con IPIV >1.2 en pollos 6 semanas de edad o causar mortalidad >75% en pollos 4-8 semanas edad inoculados vía intravenosa".
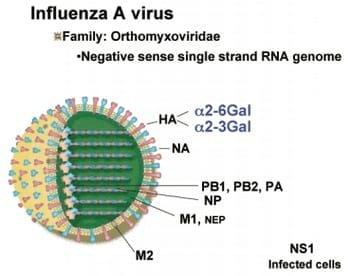
Estructura de los virus de Influenza Aviar. Nótese el genoma divido en ocho segmentos
Período de incubación
El período de incubación de los virus de IA en aves infectadas naturalemente puede ser muy corto, 24 a 36 horas, dependiendo de la virulencia del agente y de la susceptibilidad del hospedador, puede llegar hasta 14 días. Desde el punto de vista clínico se toma un promedio de 7 días.
Patogenia
El proceso infeccioso se inicia con la inhalación o ingestión de lo viriones infectivos, a continuación ocurre la colonización de las vías respiratorias altas, en particular, la penetración ocurre por la mucosa nasal y la faringe, o bien, por la celulas epiteliales de la mucosa intestinal del aparato digestivo. Las enzimas del tipo tripsina permiten la fijación de las hemoaglutininas ocurre la penetración viral, para que ocurran múltiples ciclos de replicación. Las partculas víricas se dispersan vía el sistema vascular para entonces replicarse en todo tipo de órganos parenquimatosos, encéfalo y piel. Los signos clínicos, sintomatología y la muerte ocurre por falla múltiple de los órganos. Los daños causados por los ortomixovirus son el resultado de tres procesos: 1) Replicación directa de los virus en células, tejidos y órganos. 2) Efectos indirectos debidos a la producción masiva de mediadores celulares, tales como las citocinas, e 3) isquemias provocadas por trombosis vascular.
Síntomas y lesiones
La intensidad de los signos y síntomas provocados por los virus de la influenza aviar, varían en intensidad dependiendo si se trata de agentes de baja o de alta patogenicidad. Los síntomas respiratorios son estornudos y estertor traqueal, bronquial, disnea y conjuntivitis. Los digestivos son enteritis con hemorrágias petequiales y equimóticas con una intensa diarrea de color verde.
Finalmente se abre un cuadro neurológico, que consiste en incoordinación, ataxia, y parálisis con opistótonos y epistótonos. Un signo más que revelador es la tortícolis y las contracciones tónico-clónicas.
Se trata de un cuadro neuro-entero-neurológico congestivo hemorrágico masivo. Quizá la única lesión patognomónica diferencial con la enfermedad de Newcastle es la presencia de sufuciones dérmicas que no son otra cosa que hemorragias bajo de la piel.
Replicación viral
Como es bien sabido, los virus de influenza presentan un alto rango de error durante su proceso de replicación, muy en particular, durante la trascripción de sus genomas, debido a la baja fidelidad de su polimerasa ARN1. Dicho riesgo de alto error, provoca la aparición de un fenómeno en el cual aparecen subespecies, entre las cuales muchos genotipos virales diferentes co-circulan dentro del huésped y cada subespecie de virus tiene potencialmente diferente habilidad para adaptarse al huésped2. La ventaja de la presencia de una alta tasa de error, consiste en la habilidad de los virus para adaptarse rápidamente a las nuevas condiciones ambientales.
Esto puede ocurrir, por ejemplo, durante la infección de una nueva especie de hospedador que requiere de cambios genéticos de adaptación para la replicación óptima y la transmisión del virus.
La principal desventaja que presenta una alta tasa de error, es la producción de muchos genotipos virales que pueden tener una menor capacidad de adaptación al hospedador original. La mayoría de los cambios de los nucleótidos son eventualmente eliminados del juego de genes debido a que ellos aportan, ya sea, un factor de selección neutral o negativo para la replicación continua del genotipo viral.
En algunas ocasiones, estas mutaciones neutrales o sinónimas pueden aumentar en su frecuencia cuando ellas se hayan ligadas a un cambio no sinónimo, que confiere al genotipo viral un factor de selección debido a un cambio al azar. Dichos cambios sinónimos y no sinónimos proveen marcadores de gran utilidad en el rastreo epidemiológico de los virus durante un brote3.
Para muchos virus, la tasa de cambio mutacional, ocurre a un ritmo constante y predecible, el cual ha sido frecuentemente llamado: "reloj molecular"4.
La tasa de cambio mutacional, incluye tanto a los cambios sinónimos como los no sinónimos, y ella puede ser determinada en los virus del mismo linaje, si ellos son muestreados a través del tiempo. La tasa evolutiva puede ser también determinada, por medio del examen de los cambios no sinónimos o del aminoácido, los cuales pueden ocurrir en virus del mismo linaje cuando se muestrean al paso del tiempo.
Los virus de influenza se encuentran en una gran variedad de especies, a menudo causando cuadros infecciosos severos. Sin embargo, entre los hospedadores naturales de los virus de influenza aviar, como son las aves acuáticas continentales y aves marítimas costeras migratorias, el virus se puede considerar inicialmente como avirulento5. Cuando un virus de influenza infecta a un hospedador de una especie nueva, puede replicarse causando la enfermedad ocasionalmente, y dicho virus raramente se transmitirá en la nueva especie para causar una epizootia. Por ejemplo, en el caso del virus H5N1 que provocó la "gripe" de los pollos en Hong Kong, pasó de pollos a seres humanos en al menos 18 casos causando seis defunciones durante 1997, mostró afortunadamente en aquel momento, poca capacidad para causar una epidemia de carácter mundial6.
La transmisión entre mamíferos, también puede ocurrir de manera regular, por ejemplo, existen numerosos reportes de virus de influenza porcina que han infectado a seres humanos (zoonosis) y viceversa (antropozoonosis)7. Afortunadamente, los brotes epidémicos resultantes de la transmisión de virus entre especies diferentes son raros. El cruzamientos de virus entre especies puede resultar en la introducción de un nuevo subtipo, portador de una nueva hemoaglutinina y/o neuraminidasa a los seres humanos, por medio de un cambio antigénico, el cual puede conducir a una epidemia o pandemia (caso reciente de la Influenza Pandémica A/H1N1 de marzo-abril del 2009 y que se haya aun esta en curso), debido a que el nuevo huésped no posee inmunidad protectora suficiente contra la nueva cepa recombinada de influenza. Este tipo de cambios antigénicos entre los virus de influenza han ocurrido en tres ocasiones en la población avícola de este planeta durante el siglo pasado8. Aunque las introducciones de virus de influenza en aves han sido frecuentes, estos brotes una vez que se han descubierto, usualmente, no han continuado o persistido durante largo tiempo debido a los esfuerzos por controlarlos o por la falla de los virus a adaptarse al nuevo huésped. Sin embargo, al menos han ocurrido tres brotes que se han extendido a lo largo de los años y de los cuales se han aislado múltiples virus. Estos casos han sido el brote por el subtipo H5N2 de Pennsylvania de 1983-19849, el brote ocurrido en México por un virus H5N2 de 1994-1995 hasta la fecha10, y el brote reportado en el Noreste de los Estados Unidos por el subtipo H7 en los mercados de aves vivas de 1999 hasta el presente11. Estos tres brotes han sido una estupenda y única oportunidad para poder estudiar los cambios evolutivos en las aves domésticas, primariamente en los pollos y en los pavos.
Ecología viral
Aunque los virus de la influenza de las aves pueden infectar una amplia variedad de aves y mamíferos, el hospedador natural de estos virus son las aves silvestres acuáticas migratorias.
Cuando otras especies animales, incluyendo pollos, cerdos, caballos, y seres humanos, son infectados por virus de Influenza, estos últimos pueden considerarse como hospedadores aberrantes. La distinción entre un huésped normal y uno aberrante, es de gran importancia, cuando se intenta determinar la evolución de los virus en y entre diferentes grupos de hospedadores. La tasa evolutiva de un virus de Influenza en los reservorios del hospedador natural tiende ser lenta, mientras que en los mamíferos es más rápida. La tasa de evolución mayor en mamíferos, se piensa es debida, a la presión selectiva ejercida sobre los virus para adaptarse a una especie hospedadora aberrante.
Los virus de Influenza Aviar aislados a partir de pollos y de pavos, han sido erróneamente clasificados juntos con los ortomixovirus de aves silvestres acuáticas, continentales y marítimas migratorias, cuando se determinaron las tasas de cambio evolutivo con el objeto de establecer las tasas de capacidad evolutiva y mutacional de un virus de cualquier especie hospedadora. Para esto, dos condiciones primarias deben asumirse, primera: todos los aislamientos incluidos en el estudio y análisis, deben de haber descendido de una simple y sola introducción de virus. La segunda: un brote debe permanecer el tiempo suficiente y necesario, para poder establecer y determinar una tendencia. En el caso de las aves domésticas han ocurrido recientemente tres brotes, los cuales cumplen con estos criterios, ya que la secuencia de la hemoaglutinina y de los genes no estructurales fue estudiada. Las secuencias de éstos tres virus que causaron los brotes, fueron comparados con virus de influenza, las cuales a nivel de aminoácidos son altamente conservados por todas las proteínas virales internas. Este consenso en la secuencia provee también, un punto de origen común para comparar todos los virus de Influenza. Las tasas evolutivas que se determinaron en los tres brotes, fueron similares a los que se observó en mamíferos, lo cual provee una fuerte evidencia de la capacidad de adaptación de los virus de Influenza a nuevas especies de huéspedes, como es el caso del pollo y del pavo.12
Proceso Evolutivo
El primer brote de Influenza Aviar de Alta Patogenicidad, fue reportado y descrito por el científico italiano Edoardo Perroncito en 187813. Se trata de un fascinante reporte que describe la devastadora y virulenta enfermedad como una entidad infecto-contagiosa que afectó, las aves de corral criadas en los valles y colinas alrededor de la ciudad de Turín, en el Piamonte italiano. Dicha epizootia, se inició con un cuadro clínico caracterizado por signos benignos, que subsecuentemente se transformó en un cuadro neumo-entero-encefálico severo, que mató a la casi totalidad de las aves criadas en el área mencionada. Es muy posible que los signos y síntomas inicialmente descritos por Perroncito correspondan a la forma causada por los virus de Influenza Aviar de Baja Patogenicidad y que subsecuentemente, después de la circulación dentro de la población susceptible, los virus hayan mutado a la forma de Alta Patogenicidad. De hecho, gracias a las modernas pruebas de Biología Molecular, ha sido posible demostrar que los virus de Alta Patogenicidad han mutado a partir de virus progenitores de Baja Patogenicidad de los subtipos H5 y H7, como han sido los casos de los brotes de Pennsylvania 1993-94, de México 1994-95, Italia de 1999-2000 y Chile del 2000.
La etiología viral de la IAAP, no fue identificada sino hasta 1901, por otros dos científicos italianos, Centanni y Savonuzzi, quienes demostraron que la "Peste Aviar", podía ser reproducida en el laboratorio administrando homogenizados ultrafiltrados en filtros de cerámica, obtenidos de órganos colectados de aves muertas por dicha enfermedad.14
La mayor parte de virus de IA han sido aislados de aves silvestres, particularmente de las acuáticas pertenecientes a los Órdenes Anseriformes y Charadriiformes. Es más, estas aves al parecer fungen como reservorios naturales y de hecho, funcionan como almacenes de genes, lo que permite la perpetuación de los virus de Influenza Aviar en la naturaleza. Sin embargo, es muy importante señalar que dichas aves perpetúan solamente virus de Baja Patogenicidad, ya que a pesar del enorme número de aislamientos de virus de Baja Patogenicidad que se hacen de estas especies de volátiles, muy pocas veces se han aislado virus de Alta Patogenicidad, por lo que se ha sugerido que ellas son los hospedadores naturales de los virus de Influenza Aviar, y en los que los virus se han adaptado adecuadamente.
Contrariamente, las aves domésticas, es decir, los individuos del Orden Galliformes, Familia Phaisanidae, no parecen ser los hospedadores naturales de los ortomixovirus, por lo tanto, el grado de adaptación al huésped es bajo, lo cual podría posiblemente explicar porqué las mutaciones virales documentadas han ocurrido prácticamente siempre en aves domésticas. De hecho, el único brote de Influenza Aviar de Alta Patogenicidad en aves silvestres que haya sido reportado asociado a con la mortalidad de 1,300 golondrinas de mar (Sterna hirundo) fue en 1961 en África del Sur.
Brotes de Influenza Aviar de Alta Patogenicidad reportados desde 1955
Desde que se comprobó que los brotes de IAAP son causados por virus muy virulentos de Influenza en 1955, y la demostración de que los virus de los subtipos H5 y H7, son los principales responsables de los brotes de IAAP en 1959, veinte y dos epizootias han sido reportadas en las aviculturas industriales en diferentes partes del mundo15. De éstas, veinte y una han ocurrido en aves domésticas y solamente una ha ocurrido, como hemos mencionado, en golondrinas marítimas silvestres. Tomando en cuenta la zona geográfica en donde se han reportado los brotes, en relación al desarrollo y la estructura de la avicultura del país en cuestión y en lo concerniente a las políticas implementadas de erradicación o de vacunación, las epizootias de IAAP, han tenido un impacto diferente en la economía de los países afectados. Por ejemplo, sin duda alguna, la epizootia ocurrida en la Lombardía y en el Veneto italianos y después en el resto de Italia, causada por virus de Alta Patogenicidad H7N3, entre 1999 y 2000, ha sido la más costosa que se tenga conocimiento.
Brotes documentados causados por virus altamente patogénicos de Influenza Aviar, reportados desde la identificación del agente causal de la Influenza Aviar en 1955. Modificado por Swayne y Suarez16 y Alexander.17
La mayor parte de virus de IA han sido aislados de aves silvestres, particularmente de las acuáticas pertenecientes a los Órdenes Anseriformes y Charadriiformes. Es más, estas aves al parecer fungen como reservorios naturales y de hecho, funcionan como almacenes de genes, lo que permite la perpetuación de los virus de Influenza Aviar en la naturaleza. Sin embargo, es muy importante señalar que dichas aves perpetúan solamente virus de Baja Patogenicidad, ya que a pesar del enorme número de aislamientos de virus de Baja Patogenicidad que se hacen de estas especies de volátiles, muy pocas veces se han aislado virus de Alta Patogenicidad, por lo que se ha sugerido que ellas son los hospedadores naturales de los virus de Influenza Aviar, y en los que los virus se han adaptado adecuadamente.
Contrariamente, las aves domésticas, es decir, los individuos del Orden Galliformes, Familia Phaisanidae, no parecen ser los hospedadores naturales de los ortomixovirus, por lo tanto, el grado de adaptación al huésped es bajo, lo cual podría posiblemente explicar porqué las mutaciones virales documentadas han ocurrido prácticamente siempre en aves domésticas. De hecho, el único brote de Influenza Aviar de Alta Patogenicidad en aves silvestres que haya sido reportado asociado a con la mortalidad de 1,300 golondrinas de mar (Sterna hirundo) fue en 1961 en África del Sur.
Brotes de Influenza Aviar de Alta Patogenicidad reportados desde 1955
Desde que se comprobó que los brotes de IAAP son causados por virus muy virulentos de Influenza en 1955, y la demostración de que los virus de los subtipos H5 y H7, son los principales responsables de los brotes de IAAP en 1959, veinte y dos epizootias han sido reportadas en las aviculturas industriales en diferentes partes del mundo15. De éstas, veinte y una han ocurrido en aves domésticas y solamente una ha ocurrido, como hemos mencionado, en golondrinas marítimas silvestres. Tomando en cuenta la zona geográfica en donde se han reportado los brotes, en relación al desarrollo y la estructura de la avicultura del país en cuestión y en lo concerniente a las políticas implementadas de erradicación o de vacunación, las epizootias de IAAP, han tenido un impacto diferente en la economía de los países afectados. Por ejemplo, sin duda alguna, la epizootia ocurrida en la Lombardía y en el Veneto italianos y después en el resto de Italia, causada por virus de Alta Patogenicidad H7N3, entre 1999 y 2000, ha sido la más costosa que se tenga conocimiento.
Brotes documentados causados por virus altamente patogénicos de Influenza Aviar, reportados desde la identificación del agente causal de la Influenza Aviar en 1955. Modificado por Swayne y Suarez16 y Alexander.17
| Virus de Influenza Aviar | Subtipo | País |
| A/chicken/Scotland/59 | H5N1 | Escocia |
| A/tern/South Africa/61 | H5N3 | Africa del Sur |
| A/turkey/Ontario/7732/66 | H7N3 | Canadá |
| /chicken/Victoria/76 | H7N7 | Australia |
| AA/chicken/Germany/79 | H7N7 | Alemania |
| A/turkey/England/199/79 | H7N7 | Inglaterra |
| A/chicken/Pennsylvania/1370/83 | H5N2 | EUA |
| A/turkey/Ireland/1378/83 | H5N8 | Irlanda |
| A/chicken/Victoria/85 | H7N7 | Australia |
| A/turkey/England/50-92/91 | H5N1 | Inglaterra |
| A/chicken/Victoria/92 | H7N3 | Australia |
| A/chicken/Puebla/8623-607/94 | H5N2 | México |
| A/chicken/Queretaro/14588-19/95 | H5N2 | México |
| A/chicken/Queensland/95 | H7N3 | Australia |
| A/chicken/Pakistan/447/95 | H7N3 | Pakistan |
| A/chicken/Pakistan/1369-CR2/95 | H7N3 | Pakistan |
| A/Hong Kong/220/97 | H5N1 | China |
| A/chicken/New South Wales/1651/97 | H7N4 | Australia |
| A/chicken/Italy/330/97 | H5N2 | Italia |
| A/turkey/Italy/4580/99 | H7N1 | Italia |
| A/chicken/Chile/2000 | H7N3 | Chile |
| A/chicken/Holland/2003 | H7N7 | P. Bajos, Bélgica, Alemania |
| A/chicken/Indonesia/2003 | H5N1 | Indonesia |
| A/chicken/South Korea/2003 | H5N1 | Corea del Sur |
| A/chicken/Japan/2004 | H5N1 | Japón |
| A/chicken/Taiwan/2004 | H5N2 | Taiwan |
| A/chicken/Hong Kong/2004 | H5N1 | Hong Kong |
| A/chicken/Pakistan/2004 | H7N3 | Pakistan |
| A/chicken/Viet Nam/2004 | H5N1 | Viet Nam |
| A/chicken/Thailand/2004 | H5N1 | Tailandia |
| A/chicken/Cambodia/2004 | H5N1 | Camboya |
| A/chicken/Laos/2004 | H5N1 | Laos |
| A/chicken/China/2004 | H5N1 | China |
| A/chicken/Malaysia/2004 | H5N1 | Malaysia |
| A/chicken/ North Korea/2005 | H7N1 | North Korea |
Como adición a lista arriba descrita, un gran número de aislamientos virales a partir de aves silvestres, se han dado en forma de cascada, entre 2006 y 2009 en Rusia, Ucrania, Croacia, Turquía, Irán, Irak, Grecia, Italia, Francia, España y EUA, etc., todos ellos por virus H5N1, algunos de Alta y otros de Baja Patogenicidad, así como, la mayoría de los brotes de IAAP en aves domésticas ocurridos del 2005 al segundo semestre del 2009 en Asia, Europa y África, particularmente en Indonesia, Viêt Nam y Egipto.
Además de los brotes arriba descritos, se han documentado las epizootias de Italia en 1878, Egipto en 1923 y 1945 y la de Estados Unidos en 1929. Los brotes más devastadores han sido los de Pennsylvania, con pérdidas de más de 464 millones de dólares, el de México que afectó a 25 millones de aves y el de Italia con el sacrificio de 17 millones de individuos. En Octubre del 2002, se reportó por medio de la prueba de Inhibición de la Hemoaglutinación, nueva y desalentadoramente, la presencia de un virus de Influenza Aviar de Baja Patogenicidad, en este caso un subtipo H7N3 en pavos, en las provincias de Brescia, Mantova, Vicenza y Verona en el norte Italia, con la notificación de 140 focos. El análisis filogenético llevado a cabo en el gen de la hemoaglutinina H, indicó que este aislamiento forma parte del linaje Euroasiático H7. Dicho virus esta relacionado con el subtipo H7N1, pero no es parecido al que provocó el brote epizoótico de 1999-2001 en Italia. El virus, tampoco tiene ninguna relación con el virus vacunal del subtipo H7N3, con el cual se elaboró la vacuna inactivada en aceite que se uso para el control del brote anterior (A/ck/Pakistan/95). Tan pronto como la infección parecía salirse de control, se estableció una campaña de vacunación aprobada previamente por la Comisión Veterinaria de la Comunidad Económica Europea18. Asimismo, todos estamos concientes del brote de IA de Alta Patogenicidad que ha ocurrido en marzo del año 2003 en los Países Bajos y el cual en el transcurso de semanas se extendió a las fronteras de Bélgica y después de Alemania. En los tres casos se ha procedido a la erradicación por sacrificio.
La catástrofe ocurrida a fines del año 2003 y a lo largo del 2004, 2005 y 2006 en el Sureste Asiático, causada por ortomixovirus de Alta Patogenicidad, H5N1, que afectaron inicialmente, Indonesia, Corea del Sur, Japón, Viêt Nam, Tailandia, Camboya, Laos y China y el subtipo H7N3 de Alta Patogenicidad que infectó a Pakistán, son un excelente ejemplo de la peligrosidad y rapidísima difusión actual de los patógenos, debido a los medios masivos de transportación aérea, que potencialmente pueden difundir y contagiar la población animal y/o humana en cuestión de 48-72 horas. Las pérdidas que ha sufrido la industria avícola asiática han sido cuantiosas, además de lo terriblemente preocupante que fue el escape viral zoonótico de los ortomixovirus aviares a la población humana en países como Indonesia, Viêt Nam y Egipto por mencionar los más importantes, son prueba de la aceleración de las posibilidades de mutación por recombinación (Drift) o por error en la hemoaglutinina o neuroaminidasa (Shift), la que ostentan estos virus, son indudablemente una potencial amenaza peligro para la población humana. Todo esto obligó a la Oficina Internacional des Epizootias (OIE) de Paris, a la Organización Mundial para la Agricultura y Alimentación (FAO) de Roma y al Organización Mundial de la Salud (WHO) de Ginebra, Suiza, a organizar una reunión de emergencia para el control y prevención de la Influenza Aviar en Bangkok, Tailandia, en febrero del 2004 y abril del 2005, para tomar las acciones correctivas, con el objeto de evitar una nueva gran panzootia de Influenza Aviar a nivel mundial. Es claro y evidente que la Influenza Aviar, se ha ido transformando de una enfermedad de carácter local, a regional a mundial. Otro hecho contundente es que ante la globalización de las economías del mundo, la IA se ha mundializado y que ha pasado de ser una enfermedad exótica a una entidad de carácter enzoótico/endémico en muchos países del mundo, principalmente en el Sureste Asiático.
Situación de la Influenza Aviar en el Continente Americano
Los únicos paises en América que han sido afectados recientemente por virus de baja y/o de alta patogenicidad de IA han sido los Estados Unidos en 1983-1984 por virus de IAAP H5N2, México en 1994-1995 por virus de IAAP H5N2, Canadá en 2007 y 2008 por virus IAAP H7N3, Guatemala, por virus de IABP H5N2 en el 2000 y El Salvador en el 2001, República Dominicana por virus de IABP en el 2008 y Haití en el mismo año. El resto de las otras naciones del Continente Americano, afortunadamente se han mantenido libres de la influenza aviar.
Prevención
La mejor acción para para un pais, zona o compartimento en el el control y la prevención de la influenza aviar es la erradicación total por medio del sacrificio de los animales enfermos y la destrucción de todo alimento, cama, y el lavado y desinfección de los galpones y material contaminado, de acuerdo a las normas especificadas por la OIE en su Código de Salud para los Animales Terrestres.
Vacunación
En algunos paises las caraterísticas de sus industrias avícolas y las circunstancias como se han presentado los brotes y epizootias causadas por virus de alta y de baja patogenicidad de la IA, han obligado a algunos países a recurrir a la vacunación, como han sido los casos de México, Guatemala, El Salvador, Italia, algunos estados de la Unión Americana, Tailandia, China, Indonesia, y más recientemente Egipto. Existen dos tipos de inmunógenos para la prevención de la IA, las vacunas inactivadas en vehículo oleoso y las vacunas recombinantes. Las primeras se han empleado amplia y masivamente en los paises arriba mencionados. Son muy seguras, inmunogénicas y económicas. Se aplican por vía subcutánea en la parte media posterior del cuello, ya sea a un día de edad en incubadora, pero más comunmente en granja entre los 7 y 10 días de edad de las aves. Las segundas son igualmente eficientes, se aplican a un día de edad en la incubadora, pero su costo puede ser alto.
Conclusiones
Del listado antes mencionado, podemos observar que las hemoaglutininas más comunes en patología e infectología aviar, son la H5 y la H7. Además, de los brotes de IAAP ocurridos desde 1955, el 58 % de los casos fueron causados por subtipos H5 y el 42 % de los brotes fueron provocados por subtipos H7. Todos los continentes han sido afectados, siendo los países que se hallan dentro del paralelo 60º Latitud Norte, incluyendo el brote de Escocia de 1959 y los que se hallan dentro de los 45º de Latitud Sur, considerando el brote de Chile del 2002, han sido las regiones geográficas más afectadas. Sin embargo, las epizootias que han sido más comunes han ocurrido entre los 30º, tanto de Latitud Norte, como 30º de Latitud Sur, como fue el caso de la presencia del virus H5N2 de Baja Patogenicidad en República Dominicana al inicio del 2008.
Es evidente que nos encontramos ante una preocupante reactivación y aceleración de esta ortomixovirosis, provocada sin duda alguna, por el intenso comercio intra e intercontinental, que ha favorecido la introducción de virus de Influenza Aviar Baja Patogenicidad, en aviculturas de regiones geográficas que estaban libres y que consideraban a esta infección como una enfermedad exótica. Conforme vaya aumentando la comercialización, irá por ende, aumentando el riesgo y la alta posibilidad de la aparición de un mayor número de brotes con una mayor frecuencia.
Los Departamentos de Salud Animal de los países que cuenten con una avicultura industrial de poca consideración o de gran importancia, deberán todos, implementar las medidas sanitarias estipuladas por la OIE y deberán de capacitar a personal veterinario y a profesionales afines, tales como virólogos, inmunólogos, epizootiólogos/epidemiólogos, y expertos en biología molecular, para establecer campañas de vigilancia pasiva y activa permanentemente a nivel nacional e internacional, llevar a cabo muestreos constantes, aislamiento y estudio de las secuencias de amino ácidos en el sitio de ruptura de las hemoaglutininas, con el objeto de construir los arboles filogenéticos y los linajes de los virus aislados con el objeto de poder hacer el rastreo de los mismos.
La industria avícola mundial, las naciones y los organismos internacionales, deberán levantar los fondos económicos necesarios para financiar la construcción y el funcionamiento de laboratorios a nivel nacional, regional y global y para organizarse por medio de una red global de laboratorios de diagnóstico de referencia y de investigación para el estudio de esta enfermedad, con el objeto de hacer exitosamente el control, la prevención y la erradicación de esta ortomixovirosis.
En la mayoría de los países se ha optado con éxito, por la erradicación por medio del sacrificio de cientos de miles o millones de aves, como fue el caso chileno, sin embargo, en otros países como México, Guatemala, El Salvador, Pakistán e Italia, debido a las características y circunstancias específicas y especiales que enfrentaron en su momento, se vieron obligados ha recurrir a la vacunación para el control y prevención de esta enfermedad, haciendo consecuentemente que ahora, la Influenza Aviar haya tomado, en ellos un carácter enzoótico, mientras que en otros países o regiones la Influenza Aviar sea considerada aun, como una entidad infecciosa exótica.
La industria avícola mundial, las naciones y los organismos internacionales, deberán levantar los fondos económicos necesarios para financiar la construcción y el funcionamiento de laboratorios a nivel nacional, regional y global y para organizarse por medio de una red global de laboratorios de diagnóstico de referencia y de investigación para el estudio de esta enfermedad, con el objeto de hacer exitosamente el control, la prevención y la erradicación de esta ortomixovirosis.
En la mayoría de los países se ha optado con éxito, por la erradicación por medio del sacrificio de cientos de miles o millones de aves, como fue el caso chileno, sin embargo, en otros países como México, Guatemala, El Salvador, Pakistán e Italia, debido a las características y circunstancias específicas y especiales que enfrentaron en su momento, se vieron obligados ha recurrir a la vacunación para el control y prevención de esta enfermedad, haciendo consecuentemente que ahora, la Influenza Aviar haya tomado, en ellos un carácter enzoótico, mientras que en otros países o regiones la Influenza Aviar sea considerada aun, como una entidad infecciosa exótica.
Referencias Bibliográficas
1 Parvin, J.D., A. Moscona, W.T. Pan, J.M. Leider, P. Palese. 1986. Measurement of the mutation rates of animal viruses: influenza A virus and poliovirus type 1. J. Virol. 59, 377-383.
2 Domingo, E., E. Martínez Salas, F. Sobrino, J.C. de la Torre, A. Portela, C. López Galidez, P. Pérez-Brena and Villanueva and R. Nájera. 1985. The quasispecies (extremely heterogeneous) nature of viral RNA genome populations: biological relevance- a review. Gene 40, 1-8.
3 Lindstrom, S., A.Endo, S. Sugita, M. Pecoraro, Y. Hiromoto, M. Kamada, T. Takashahi, K. Nerome, 1998. Phylogenetic analysis of the matrix and non-structural genes of equine influenza viruses. Arch. Virol. 143, 1585-1598
4 Buonaugurio, D. A., S. Nakada, J.D. Parvin, M. Krystal, P. Palese, W.M. Fitch. 1986. Evolution of human influenza viruses over 50 years, uniform rate of change in NS gene. Science 232, 980-982.
5 Slemons, R.D., D.C. Johnson, J.S. Osborn, F. Hayes, 1974. Type-A influenza viruses isolated from wild free-flying ducks in California. Avian Dis. 18, 119-124.
6 Mounts, A.W., H. Kwong, D.S. Izurieta, Y.Y. Ho, T.K. Au, M. Lee, C. Buxton-Bridges, S.W. Williams, K.H. Mak, J.M. Katz, W.W. Thompson, N.J. Cox, K. Fukuda, 1999. Case-control study of risk factors for avian influenza A (H5N2) disease, Hong Kong, 1997. J. Infect. Dis. 180, 505-508.
7 Dowle, W.R., A.W. Hattwick, 1977. Swine influenza virus infections in humans. J. Infect. Dis. 136, S386-S389.
8 Webster, R.G., W.J. Bean, O.T. Gorman, T.M. Chambers, Y. Kawaoka, 1992. Evolution and ecology of influenza viruses. Microbiol. Rev. 56, 152-179.
9 Suarez, D. L., D.A. Senne, 2000. The link between the live bird markets and the Pennsylvania H5N2 avian influenza outbreak of 1983-1984. Avian Dis.
10 Garcia, M., D.L. Suarez, J.M. Crawford, J.W. Latimer, R.D. Slemons, D.E. Swayne, M.L. Perdue, 1997. Evolution of H5 subtype avian influenza viruses in North America. Virus Res. 51, 115-124.
11 D.L., M. Garcia, J. Latimer, D. Senne, M. Perdue, 1999. Phylogenetic analisis of H7 avian influenza viruses isolated from the live bird markets of the Northeast United States. J. Virol. 73, 3567-3573.
12 Suarez, David L. Evolution of avian influenza viruses. Veterinary Microbiology. 74 (2000). 15-27.
13 Perroncito, E. Epizzotia tifoide nei gallinacei. Annali Accademia Agricoltura. Torino. 21, 87-126. 1878.
14 Centanni E., E. Savunozzi: La Peste Aviaria. Atti dell’Accademia delle Scienze Mediche e Naturalli di Ferrara. 111-123, 1901.
15 Alexander, D.J.: The history of avian influenza in poultry. World Poultry No. Special, 7-8, 2000.
16 Swayne D.E. and D.L. Suarez. Highly pathogenic avian influenza. Rev. Sci. Tech. Off. Int. Epiz. 19, 463-482.
17 Alexander D.J. A review of avian influenza in different bird species. Vet Microbiol. 74, 3-13, 2000.
16 Swayne D.E. and D.L. Suarez. Highly pathogenic avian influenza. Rev. Sci. Tech. Off. Int. Epiz. 19, 463-482.
17 Alexander D.J. A review of avian influenza in different bird species. Vet Microbiol. 74, 3-13, 2000.
18 Capua, Ilaria, Stefano Maragnon, Calogero Terregino and Giovani Cattoli: The Avian Influenza Experience, AI in Italy, 1997-2002. Lecture given during the Educational Program of the 55th International Poultry Exposition. January 23, 2003. Atlanta, Georgia. USA.
Temas relacionados
Autores:

Únete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicación



Te puede interesar
.jpg&w=3840&q=75)