Enzimas fibrolíticas exógenas en la digestibilidad in vitro de cinco ecotipos de Brachiaria
Publicado: 17 de octubre de 2007
Por: Juan H. Avellaneda-Cevallos, Sergio S. González-Muñoz, Juan M. Pinos-Rodríguez, Alfonso Hernández-Garay, Oziel D. Montañez-Valdez, José Ayala-Oseguera
Los forrajes representan la fuente más económica para la alimentación de los rumiantes, principalmente en el trópico donde hay grandes extensiones de tierra dedicadas a la explotación ganadera. En casi todos los países tropicales la expansión de la frontera agrícola llegó a su límite, y el crecimiento actual de la actividad agropecuaria depende en alto grado de la intensificación y tecnificación de la producción.
En las zonas tropicales de México se produjeron 1.333.100.000 litros de leche en el año 2000 (SAGARPA 2000) y 542.724 t de carne en el 2001 (Gallardo et al. 2002). El hato ganadero en estas zonas se alimenta principalmente mediante pastoreo en áreas cubiertas por grama nativa y especies introducidas (Gallardo et al. 2002). Por tal razón, en estos ecosistemas es importante que los ganaderos dispongan de opciones forrajeras que aumenten la productividad pecuaria, con un enfoque sostenible. Esto permitirá sustituir o complementar especies poco productivas por otras de mayor rendimiento, y con mejores características agronómicas, con las cuales será posible una producción sostenible de la pradera y del hato ganadero.
En este sentido, el género Brachiaria tiene gran potencial como recurso genético forrajero, ya que tiene gran capacidad de adaptación a una am plia ga ma de condiciones edáficas y climáticas (Valle 1990, citado por En rí quez y Ro me ro 1999). En la dé ca da de 1980, el Instituto de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) - México, evaluó varios ecotipos del género Brachiaria de los cua les tres fue ron for malmente liberados: B. decumbens var. Se ñal o Chon tal po, B. brizantha var. Insur gente y B. humidicola var. Chetu mal, los cua les re sultaron sobresalientes en diversos ecosistemas del trópico mexicano (Enríquez y Pe ralta 1992, ci ta dos por En rí quez y Ro me ro 1999).
Sin embargo, las gramíneas tienen una alta concentración de pared celular (65 – 75 %) y baja digestibilidad (45 – 55 %), factores asociados con una reducción en consumo voluntario y menor calidad nutritiva del forraje (Gallean y Goetsch 1993). Por tal motivo, el objetivo de este experimento fue evaluar el efecto de un compuesto enzimático fibrolítico exógeno en la degradación in vitro de la materia seca y fracciones de la pared celular de cinco ecotipos del género Brachiaria.
MATERIALES Y MÉTODOS
Esta investigación se realizó en el laboratorio de bromatología del Colegio de Postgraduados ubicado en Montecillo, Estado de México, México entre el 25 de abril y 27 de no viem bre del 2002. Se uti li za ron cua tro bo rre gos Suffolk (74±2 kg PV) con cá nu la ru mi nal y alimentados ad libitum (a libertad) con he no de pas to guinea (Panicum maximum var. Mombasa), premezcla mineral y agua, en la unidad metabólica para rumiantes, del Pro gra ma de Ga na de ría, de la gran ja experimental del Colegio de Postgraduados, Montecillo, Estado de México, México. Se usa ron he nos de cin co eco ti pos de Brachiaria [(brizantha var. To ledo (BT), ruziziensis x decumbens CIAT 46024 (RD); decumbens var. Se ñal (DS); ruziziensis x brizantha CIAT 36061 cv. Mu la to (RBM); brizantha var. Insur gente (BI)] cor ta do a una edad de 35 d y he ni fi ca do a la in tem pe rie en Ciu dad Isla, Estado de Ve racruz, México, entre el 14 de octubre de 2001 y 11 de ene ro de 2002.
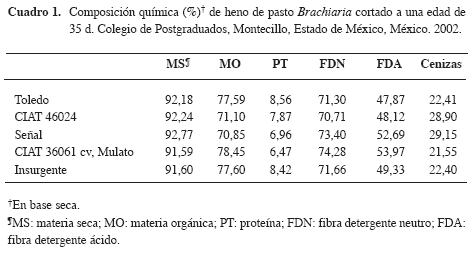
El compuesto enzimático fibrolítico exógeno (Fibrozyme®, Alltech INC, Nicholasville, KY, U.S.A) es una com bi na ción de ex trac to de la fer men ta ción de Asper gillus ní ger y Trichoderma viride y fer men tos solubles, protegidos por técnicas de glu cosilación. Su actividad xilanásica es de 100 UI/g (una uni dad xilanásica es la cantidad de enzima necesaria para liberar 1 μmol de xi lo sa). En los he nos se mi dió ma te ria se ca (MS) y or gá ni ca (MO), pro teí na to tal (PT; N x 6,25) y ce ni zas (AOAC 1990); fi bra de ter gen te neu tro (FDN) y áci do (FDA) (Van Soest et al. 1991) (Cuadro 1).
Para la digestibilidad in vitro (DIV) de la MS (DIVMS) se utilizó la primera fase de la técnica de Tilley y Terry (1963), mezclando saliva artificial (McDougall 1948) y líquido ruminal (relación 4:1).
Con una bomba de vacío, se tomó un litro de líquido ruminal de cada borrego, se mezcló, se filtró con cuatro capas de gasa y depositó en un termo (39 °C) para ser gaseado con CO2.
El medio (40 ml saliva de McDougall y 10 ml de líquido ruminal) se depositó en tubos de propileno (100 ml) con 500 mg de heno de pasto Brachiaria molido (malla de 1 mm) y secado a 60 °C durante 24 h, sin y con 1,5 g compuesto enzimático/kg MS de Brachiaria. Los tubos fueron gaseados con CO2, tapados para mezclar su contenido y colocados en baño María (39 °C); la incubación se hizo durante 3, 6, 12, 24, 48 y 72 h y fue detenida por congelación (0 °C). Los tubos fueron descongelados a temperatura ambiente para filtrar su contenido a través de papel filtro 541 (Ø poro 10 μm) y luego secados a 60 °C para medir la DIVMS. En las fracciones de FDN y FDA, la digestibilidad in vitro (DIVFDN y DIVFDA) se determinó según Tilley y Terry (1963).
El diseño experimental fue completamente al azar con un arreglo factorial 5x2 (cinco ecotipos de Brachiaria; compuesto enzimático 0 y 1,5 g/kg MS de Brachiaria) en los seis tiempos de incubación (3, 6, 12, 24, 48, 72 h). Se usaron tres tubos (repeticiones) para cada ecotipo y la prueba de digestibilidad se hizo en dos corridas, para utilizar cada tubo como submuestra e incluir la interacción tratamiento x corrida como término de error en el modelo (Pinos-Rodríguez et al. 2002a). El modelo estadístico utilizado fue el siguiente:
Yijkl = μ + Ci + Aj +Bk + ABjk + εijkl
Donde:
Yijkl = Valor de la variable respuesta en la corrida i, ecotipo de Brachiaria j, g del compuesto enzimático k.
μ = Media general
Ci = Efecto de la corrida i
Aj= Efecto del ecotipo de Brachiaria j
Bk = Efecto de g del compuesto enzimático k
ABjk = Efecto de la interacción ecotipo de Brachiaria
j y g del compuesto enzimático k
εijkl= Error experimental
Los datos se analizaron usando el procedimiento GLM del paquete estadístico SAS (1999) y las diferencias de medias fueron comparadas usando la prueba de Tukey (p<0,05).
Para ver la imagen en tamaño real, click aquí
RESULTADOS Y DISCUSIÓN
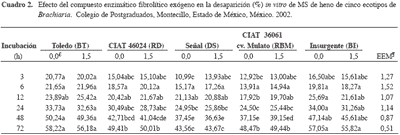
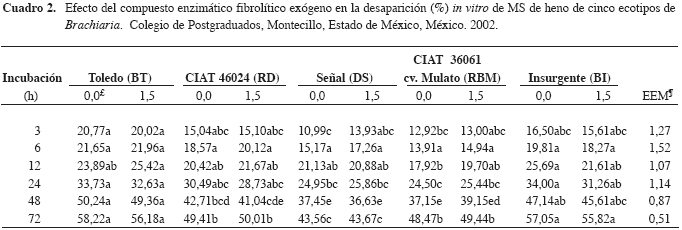
La DIVMS no fue diferente (p>0,05) entre los ecotipos durante las primeras 24 h de incubación (Cuadro 2), probablemente porque ese tiempo no es suficiente para cambiar esta variable u observar posibles cambios. Este resultado concuerda con Avellaneda-Cevallos et al. (2003), quienes no encontraron diferencias en la DIVMS de heno de pasto guinea (Panicum maximum var. Mombasa) de 35 y 90 d durante las primeras 48 h de incubación.
La DIVMS de los henos de los ecotipos DS y RBM fue menor (p<0,05) respecto a BT y BI a las 48 y 72 h; mientras que la DIVMS de RD y RBM fue más alta (p<0,05) que la de DS a las 72 h. Estos resultados concuerdan con Herrero et al. (2001) quienes reportaron diferencias en la DIVMS entre varios ecotipos de Brachiaria, aunque B. ruziziensis es más digerible, seguida por B. decumbens, mientras que B. brizantha y B. humidicola tiene la menor digestibilidad. Esto último difiere con lo observado en la presente investigación, ya que la digestibilidad de los henos de los ecotipos de bri zant ha pre sen ta ron me jor (p<0,05) di ges ti bi li dad que los de B. de cumbens.
Tal di fe ren cia pue de de ber se a fac to res co mo ca rac te rís ti cas agro nó mi cas de los eco ti pos, efec tos cli má ti cos, re co lec ción de las mues tras y su ma ne jo. La DIVMS pa ra el he no del eco ti po DS fue me nor (p<0,05) en 23,7; 12,3; 10,91 y 22,7 %, cuan do se compa ró con BT, RD, RBM y BI.
El compuesto enzimático no mejoró (p>0,05) la DIVMS de los ecotipos de Brachiaria (Cuadro 2).
Para ver la imagen en tamaño real, click aquí
De manera similar, el tratamiento con un compuesto enzimático no cambió la DIVMS en heno de trigo a las 24 h (Pollard et al. 2001), en heno de pasto guinea (Panicum maximum var. Mombasa) de 3 a 72 h (Avellaneda-Cevallos et al. 2003), en festuca (Festuca arundinacea) después de 18 h (Tricarico et al. 1998), en heno de alfalfa (Medicago sativa) de 0 a 36 h (Titi et al. 1998), ni en heno de pasto ballico (Lolium perenne) a las 48 y 72 h, y de alfalfa a las 24 y 72 h (Pinos et al. 2001). Sin embargo, el uso de un compuesto enzimático ha mejorado la DIVMS de heno de alfalfa (Pollard et al. 2001) y de fes tu ca en las primeras 12h (Tricaricoet al. 1998); además, según Pinosetal. (2001), aumentó la DIV de la MO del heno de ballico (48h) y de alfalfa (24, 48 y 72 h). En este caso, las diferencias en los resultados puede de berse a factores como tipo y manejo de las muestras de forraje, así como técnicas y equipo de la borato rio em pleados para medir la digestibilidad invitro.
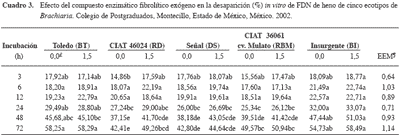
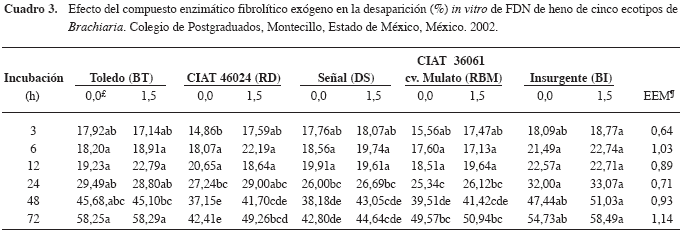
La DIVFDN (Cuadro 3) a las 48 y 72 h para los henos de los ecotipos BT y BI fue mayor (p<0,05) respecto a RD, DS y RBM; además, el compuesto enzimático aumentó (p<0,05) la DIVFDN a las 72 h del heno de RD. Lo anterior se puede deber, en parte, a diferencias entre componentes de la pared celular de los ecotipos, y porque las enzimas del compuesto requieren varias horas para cambiar la digestibilidad de la FDN. En los demás tiempos y ecotipos no hubo diferencia (p>0,05) entre el testigo (sin compuesto enzimático) y la adición de 1,5 g del compuesto enzimático (Cuadro 3). Al respecto, Avellaneda- Cevallos et al. (2003) observaron incrementos en la DIVFDN (p<0,05) a las 3, 6 y 12 h sólo para heno de guinea (Panicum maximum var. Mombasa) de 90 d, aunque a las 24, 48 y 72 h la DIVFDN fue mayor (p<0,05) para guinea de 35 d; además, la enzima (1,5 g) aumentó (p<0,05) la DIVFDN para guinea de 35 d (a las 48 h) y de 90 d (a las 72 h). Man deb vu et al. (1999) en con tra ron que la DIVFDN fue ma yor (p<0,05) pa ra un pas to ber mu da (Cy no don dacty lon) co se cha do ca da 21 d com pa ra do con otro co se cha do ca da 42 d, pe ro no hubo efecto de un compuesto en zimático exógeno. Sin embargo, según Pinos et al. (2002b), el com puesto en zi má ti co in cre men tó (p<0,05) la DIVFDN de he no de al fal fa y de pas to ba lli co.
Para ver la imagen en tamaño real, click aquí
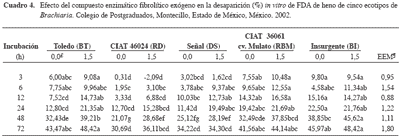
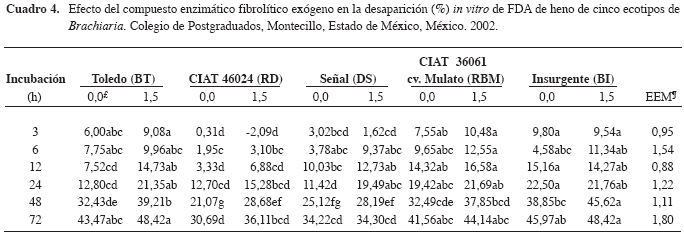
La DIVFDA (Cuadro 4) fue mayor (p<0,05) a las tres horas para los henos de los ecotipos BT, RBM y BI comparados con el RD. Además, el compuesto enzimático aumentó (p<0,05) la DIVFDA para BT (12, 24 y 48 h), DS (24 h), RBM (48 h), BI (48 h). Al respecto, la DIVFDA de heno de guinea de 35 d, a las 3, 6, 12, 24, 48 y 72, y de guinea de 90 d a las 24 y 72 h fue mayor (p<0,05) cuando se usó el compuesto enzimático (Avellaneda-Cevallos et al. 2003). No obstante, Pinos- Rodríguez et al. (2002b) no observaron diferencias en la digestibilidad in vi tro de la FDA de he no de al fal fa y de pasto ballico, al agregar un compuesto enzimático fibrolítico exógeno. La diferencia entre los resultados de este ex perimento y los publicados por otros investigadores puede deberse al tipo de forraje.
CONCLUSIONES
Con base a las condiciones experimentales descritas, se con clu ye que los he nos de los eco ti pos de B. brizantha pre sen ta ron me jor DIVMS (24, 48 y 72 h) y DIVFDN (48 y 72 h) com pa ra dos con los de B. decumbens.
El compuesto enzimático aumentó la DIVFDN para FDN a las 72 h del heno de ruziziensis x decumbens CIAT 46024 (RD); en los demás tiempos y ecotipos no hubo cambios en la DIVFDN.
La DIVFDA no fue diferente entre los ecotipos a las 6, 12, 24, 48 y 72 h. Sin embargo, el compuesto enzimático incrementó la DIVFDA de los ecotipos B. brizantha var. Toledo (BT; 12, 24 y 48 h), B. decumbens var. Señal (DS; 24 h), B. ruziziensis x B. brizantha CIAT 36061 cv. Mulato (RBM; (48 h), B. brizantha var. Insurgente (BI: 48 h).
De acuerdo con estos resultados experimentales, el compuesto enzimático fibrolítico exógeno empleado tuvo efectos positivos en la degradación in vitro de algunas fracciones de la pared celular, principalmente de la FDA, en los henos de los cinco ecotipos de pasto Brachiaria.
LITERATURA CITADA
En las zonas tropicales de México se produjeron 1.333.100.000 litros de leche en el año 2000 (SAGARPA 2000) y 542.724 t de carne en el 2001 (Gallardo et al. 2002). El hato ganadero en estas zonas se alimenta principalmente mediante pastoreo en áreas cubiertas por grama nativa y especies introducidas (Gallardo et al. 2002). Por tal razón, en estos ecosistemas es importante que los ganaderos dispongan de opciones forrajeras que aumenten la productividad pecuaria, con un enfoque sostenible. Esto permitirá sustituir o complementar especies poco productivas por otras de mayor rendimiento, y con mejores características agronómicas, con las cuales será posible una producción sostenible de la pradera y del hato ganadero.
En este sentido, el género Brachiaria tiene gran potencial como recurso genético forrajero, ya que tiene gran capacidad de adaptación a una am plia ga ma de condiciones edáficas y climáticas (Valle 1990, citado por En rí quez y Ro me ro 1999). En la dé ca da de 1980, el Instituto de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) - México, evaluó varios ecotipos del género Brachiaria de los cua les tres fue ron for malmente liberados: B. decumbens var. Se ñal o Chon tal po, B. brizantha var. Insur gente y B. humidicola var. Chetu mal, los cua les re sultaron sobresalientes en diversos ecosistemas del trópico mexicano (Enríquez y Pe ralta 1992, ci ta dos por En rí quez y Ro me ro 1999).
Sin embargo, las gramíneas tienen una alta concentración de pared celular (65 – 75 %) y baja digestibilidad (45 – 55 %), factores asociados con una reducción en consumo voluntario y menor calidad nutritiva del forraje (Gallean y Goetsch 1993). Por tal motivo, el objetivo de este experimento fue evaluar el efecto de un compuesto enzimático fibrolítico exógeno en la degradación in vitro de la materia seca y fracciones de la pared celular de cinco ecotipos del género Brachiaria.
MATERIALES Y MÉTODOS
Esta investigación se realizó en el laboratorio de bromatología del Colegio de Postgraduados ubicado en Montecillo, Estado de México, México entre el 25 de abril y 27 de no viem bre del 2002. Se uti li za ron cua tro bo rre gos Suffolk (74±2 kg PV) con cá nu la ru mi nal y alimentados ad libitum (a libertad) con he no de pas to guinea (Panicum maximum var. Mombasa), premezcla mineral y agua, en la unidad metabólica para rumiantes, del Pro gra ma de Ga na de ría, de la gran ja experimental del Colegio de Postgraduados, Montecillo, Estado de México, México. Se usa ron he nos de cin co eco ti pos de Brachiaria [(brizantha var. To ledo (BT), ruziziensis x decumbens CIAT 46024 (RD); decumbens var. Se ñal (DS); ruziziensis x brizantha CIAT 36061 cv. Mu la to (RBM); brizantha var. Insur gente (BI)] cor ta do a una edad de 35 d y he ni fi ca do a la in tem pe rie en Ciu dad Isla, Estado de Ve racruz, México, entre el 14 de octubre de 2001 y 11 de ene ro de 2002.
El compuesto enzimático fibrolítico exógeno (Fibrozyme®, Alltech INC, Nicholasville, KY, U.S.A) es una com bi na ción de ex trac to de la fer men ta ción de Asper gillus ní ger y Trichoderma viride y fer men tos solubles, protegidos por técnicas de glu cosilación. Su actividad xilanásica es de 100 UI/g (una uni dad xilanásica es la cantidad de enzima necesaria para liberar 1 μmol de xi lo sa). En los he nos se mi dió ma te ria se ca (MS) y or gá ni ca (MO), pro teí na to tal (PT; N x 6,25) y ce ni zas (AOAC 1990); fi bra de ter gen te neu tro (FDN) y áci do (FDA) (Van Soest et al. 1991) (Cuadro 1).
Para la digestibilidad in vitro (DIV) de la MS (DIVMS) se utilizó la primera fase de la técnica de Tilley y Terry (1963), mezclando saliva artificial (McDougall 1948) y líquido ruminal (relación 4:1).
Con una bomba de vacío, se tomó un litro de líquido ruminal de cada borrego, se mezcló, se filtró con cuatro capas de gasa y depositó en un termo (39 °C) para ser gaseado con CO2.
El medio (40 ml saliva de McDougall y 10 ml de líquido ruminal) se depositó en tubos de propileno (100 ml) con 500 mg de heno de pasto Brachiaria molido (malla de 1 mm) y secado a 60 °C durante 24 h, sin y con 1,5 g compuesto enzimático/kg MS de Brachiaria. Los tubos fueron gaseados con CO2, tapados para mezclar su contenido y colocados en baño María (39 °C); la incubación se hizo durante 3, 6, 12, 24, 48 y 72 h y fue detenida por congelación (0 °C). Los tubos fueron descongelados a temperatura ambiente para filtrar su contenido a través de papel filtro 541 (Ø poro 10 μm) y luego secados a 60 °C para medir la DIVMS. En las fracciones de FDN y FDA, la digestibilidad in vitro (DIVFDN y DIVFDA) se determinó según Tilley y Terry (1963).
El diseño experimental fue completamente al azar con un arreglo factorial 5x2 (cinco ecotipos de Brachiaria; compuesto enzimático 0 y 1,5 g/kg MS de Brachiaria) en los seis tiempos de incubación (3, 6, 12, 24, 48, 72 h). Se usaron tres tubos (repeticiones) para cada ecotipo y la prueba de digestibilidad se hizo en dos corridas, para utilizar cada tubo como submuestra e incluir la interacción tratamiento x corrida como término de error en el modelo (Pinos-Rodríguez et al. 2002a). El modelo estadístico utilizado fue el siguiente:
Yijkl = μ + Ci + Aj +Bk + ABjk + εijkl
Donde:
Yijkl = Valor de la variable respuesta en la corrida i, ecotipo de Brachiaria j, g del compuesto enzimático k.
μ = Media general
Ci = Efecto de la corrida i
Aj= Efecto del ecotipo de Brachiaria j
Bk = Efecto de g del compuesto enzimático k
ABjk = Efecto de la interacción ecotipo de Brachiaria
j y g del compuesto enzimático k
εijkl= Error experimental
Los datos se analizaron usando el procedimiento GLM del paquete estadístico SAS (1999) y las diferencias de medias fueron comparadas usando la prueba de Tukey (p<0,05).
Para ver la imagen en tamaño real, click aquí
RESULTADOS Y DISCUSIÓN
La DIVMS no fue diferente (p>0,05) entre los ecotipos durante las primeras 24 h de incubación (Cuadro 2), probablemente porque ese tiempo no es suficiente para cambiar esta variable u observar posibles cambios. Este resultado concuerda con Avellaneda-Cevallos et al. (2003), quienes no encontraron diferencias en la DIVMS de heno de pasto guinea (Panicum maximum var. Mombasa) de 35 y 90 d durante las primeras 48 h de incubación.
La DIVMS de los henos de los ecotipos DS y RBM fue menor (p<0,05) respecto a BT y BI a las 48 y 72 h; mientras que la DIVMS de RD y RBM fue más alta (p<0,05) que la de DS a las 72 h. Estos resultados concuerdan con Herrero et al. (2001) quienes reportaron diferencias en la DIVMS entre varios ecotipos de Brachiaria, aunque B. ruziziensis es más digerible, seguida por B. decumbens, mientras que B. brizantha y B. humidicola tiene la menor digestibilidad. Esto último difiere con lo observado en la presente investigación, ya que la digestibilidad de los henos de los ecotipos de bri zant ha pre sen ta ron me jor (p<0,05) di ges ti bi li dad que los de B. de cumbens.
Tal di fe ren cia pue de de ber se a fac to res co mo ca rac te rís ti cas agro nó mi cas de los eco ti pos, efec tos cli má ti cos, re co lec ción de las mues tras y su ma ne jo. La DIVMS pa ra el he no del eco ti po DS fue me nor (p<0,05) en 23,7; 12,3; 10,91 y 22,7 %, cuan do se compa ró con BT, RD, RBM y BI.
El compuesto enzimático no mejoró (p>0,05) la DIVMS de los ecotipos de Brachiaria (Cuadro 2).
Para ver la imagen en tamaño real, click aquí
De manera similar, el tratamiento con un compuesto enzimático no cambió la DIVMS en heno de trigo a las 24 h (Pollard et al. 2001), en heno de pasto guinea (Panicum maximum var. Mombasa) de 3 a 72 h (Avellaneda-Cevallos et al. 2003), en festuca (Festuca arundinacea) después de 18 h (Tricarico et al. 1998), en heno de alfalfa (Medicago sativa) de 0 a 36 h (Titi et al. 1998), ni en heno de pasto ballico (Lolium perenne) a las 48 y 72 h, y de alfalfa a las 24 y 72 h (Pinos et al. 2001). Sin embargo, el uso de un compuesto enzimático ha mejorado la DIVMS de heno de alfalfa (Pollard et al. 2001) y de fes tu ca en las primeras 12h (Tricaricoet al. 1998); además, según Pinosetal. (2001), aumentó la DIV de la MO del heno de ballico (48h) y de alfalfa (24, 48 y 72 h). En este caso, las diferencias en los resultados puede de berse a factores como tipo y manejo de las muestras de forraje, así como técnicas y equipo de la borato rio em pleados para medir la digestibilidad invitro.
La DIVFDN (Cuadro 3) a las 48 y 72 h para los henos de los ecotipos BT y BI fue mayor (p<0,05) respecto a RD, DS y RBM; además, el compuesto enzimático aumentó (p<0,05) la DIVFDN a las 72 h del heno de RD. Lo anterior se puede deber, en parte, a diferencias entre componentes de la pared celular de los ecotipos, y porque las enzimas del compuesto requieren varias horas para cambiar la digestibilidad de la FDN. En los demás tiempos y ecotipos no hubo diferencia (p>0,05) entre el testigo (sin compuesto enzimático) y la adición de 1,5 g del compuesto enzimático (Cuadro 3). Al respecto, Avellaneda- Cevallos et al. (2003) observaron incrementos en la DIVFDN (p<0,05) a las 3, 6 y 12 h sólo para heno de guinea (Panicum maximum var. Mombasa) de 90 d, aunque a las 24, 48 y 72 h la DIVFDN fue mayor (p<0,05) para guinea de 35 d; además, la enzima (1,5 g) aumentó (p<0,05) la DIVFDN para guinea de 35 d (a las 48 h) y de 90 d (a las 72 h). Man deb vu et al. (1999) en con tra ron que la DIVFDN fue ma yor (p<0,05) pa ra un pas to ber mu da (Cy no don dacty lon) co se cha do ca da 21 d com pa ra do con otro co se cha do ca da 42 d, pe ro no hubo efecto de un compuesto en zimático exógeno. Sin embargo, según Pinos et al. (2002b), el com puesto en zi má ti co in cre men tó (p<0,05) la DIVFDN de he no de al fal fa y de pas to ba lli co.
Para ver la imagen en tamaño real, click aquí
La DIVFDA (Cuadro 4) fue mayor (p<0,05) a las tres horas para los henos de los ecotipos BT, RBM y BI comparados con el RD. Además, el compuesto enzimático aumentó (p<0,05) la DIVFDA para BT (12, 24 y 48 h), DS (24 h), RBM (48 h), BI (48 h). Al respecto, la DIVFDA de heno de guinea de 35 d, a las 3, 6, 12, 24, 48 y 72, y de guinea de 90 d a las 24 y 72 h fue mayor (p<0,05) cuando se usó el compuesto enzimático (Avellaneda-Cevallos et al. 2003). No obstante, Pinos- Rodríguez et al. (2002b) no observaron diferencias en la digestibilidad in vi tro de la FDA de he no de al fal fa y de pasto ballico, al agregar un compuesto enzimático fibrolítico exógeno. La diferencia entre los resultados de este ex perimento y los publicados por otros investigadores puede deberse al tipo de forraje.
CONCLUSIONES
Con base a las condiciones experimentales descritas, se con clu ye que los he nos de los eco ti pos de B. brizantha pre sen ta ron me jor DIVMS (24, 48 y 72 h) y DIVFDN (48 y 72 h) com pa ra dos con los de B. decumbens.
El compuesto enzimático aumentó la DIVFDN para FDN a las 72 h del heno de ruziziensis x decumbens CIAT 46024 (RD); en los demás tiempos y ecotipos no hubo cambios en la DIVFDN.
La DIVFDA no fue diferente entre los ecotipos a las 6, 12, 24, 48 y 72 h. Sin embargo, el compuesto enzimático incrementó la DIVFDA de los ecotipos B. brizantha var. Toledo (BT; 12, 24 y 48 h), B. decumbens var. Señal (DS; 24 h), B. ruziziensis x B. brizantha CIAT 36061 cv. Mulato (RBM; (48 h), B. brizantha var. Insurgente (BI: 48 h).
De acuerdo con estos resultados experimentales, el compuesto enzimático fibrolítico exógeno empleado tuvo efectos positivos en la degradación in vitro de algunas fracciones de la pared celular, principalmente de la FDA, en los henos de los cinco ecotipos de pasto Brachiaria.
LITERATURA CITADA
AOAC (Association of Official Analytical Chemists). 1990. Official Methods of Analysis (15 ed.). Washington, DC. 1298 p.
AVELLANEDA-CEVALLOS, J.H.; GONZÁLEZ, S.S.; PINO-RODRÍGUEZ, J.M.; HERNÁNDEZ, A.; BARCENA, R.; COBOS, M.; HERNÁNDEZSÁNCHEZ, D.; MONTANEZ-VALDEZ, O. 2003. Effect of exogenous fibrolytic enzymes (Fibrozyme) on dry matter and cell wall in vitro digestibility of guinea grass (Panicum maximum var. Mombasa) hay. J. Animal Sci (Suppl. 1):334 (Abstr.).
ENRÍQUEZ, J. F.; ROMERO, J. 1999. Tasa de crecimiento estacional a diferentes edades de rebrote de 16 ecotipos de Brachiarias spp. en Isla, Veracruz. Agrociencia. 33:141-148.
GALLARDO, N.J.L.; GARCÍA, C.M.; ALBARRAN, M.; OCHOA, R.; ORTEGA, C. 2002. Situación actual de la producción de carne de bovino en México. Claridades Agropec. 109:3-28.
GALYEAN, M. L.; GOETSCH, A. L. 1993. Utilization of forage fiber by ruminants. In: H. Jung, D. Buxton, R. Hatfield and J. Ralph. eds. Forage Cell Wall Structure and Digestibility. American Society of Agronomy, Crop Science of America, Soil Science Society of America, Madison, Wisconsin, USA. p. 33-71.
HERRERO, M.; DO VALLE, C.B.; HUGHES, N.R.G.; SABATEL, V.; JESSOP, N.S. 2001. Measurements of physical strength and their relationship to the chemical composition of four species of Brachiaria. Anim. Feed Sci. Tech. 92:149-158.
MANDEBVU, P.; WEST, J.W.; FROETSCHEL, M.A.; HATFIELD, R.D.; GATES, R.N.; HILL, G.M. 1999. Effect of enzymes or microbial treatment of Bermudagrass forages before ensiling on cell wall composition, end products of silage fermentation and in situ digestion kinetics. Anim. Feed Sci. Tech. 77:317-329.
McDOUGALL, E.I. 1948. Studies on ruminant saliva. I. The composition and output of sheep´s saliva. Biochem. J. 70:99-109.
PINOS, J.M.; GONZÁLEZ, S. S.; MENDOZA, G.; BÁRCENA, R.; COBOS, M. 2001. Efecto de enzimas fibrolíticas glucosiladas en la digestibilidad in vitro de MS y MO de alfalfa (Medicago sativa) y ballico (Lolium perenne). Rev. Fac. Agro. (LUZ) 18:505-509
PINOS-RODRÍGUEZ, J.M.; GONZÁLEZ, S. S.; MENDOZA, G. D.; BÁRCENA, R.; COBOS, M. 2002a. Efecto de enzimas fibrolíticas exógenas en la digestibilidad in vitro de la pared celular de heno de alfalfa (Medicago sativa) o de ballico (Lolium perenne). Interciencia. 27:28-32.
PINOS-RODRÍGUEZ, J.M.; GONZÁLEZ, S. S.; MENDOZA, G. D.; MARTÍNEZ, A. 2002b. Análisis estadístico de experimentos de digestibilidad in vitro con forrajes. Interciencia. 27:143-146.
POLLARD, G.; WRIGHT, W.; BRAMBL, T.; RICHARDSON, C.; COBB, C. 2001. Effects of liquid feed supplementation and (or) cellulolytic enzymes on dry matter disappearance of either legume or grass hay. J. Dairy Sci. 84 (Suppl. 1):37 (Abstr).
SAGARPA. 2000. Situación actual y perspectivas de la producción de leche de ganado bovino en México 1990-2000. 87 p.
SAS (Statistical Analysis System). 1999. User´s guide: Statistics [CD-ROM Computer file]. North Carolina, USA. Version 8. SAS Inst. Inc., Cary, NC.
TILLEY, J.M.; TERRY, R. A. 1963. A two-stage technique for the in vitro digestion of forage crops. J. Brit. Grass. Soc. 28:104-111.
TITI, H.H.; RICHARDSON, C.R.; COBB, C.W. 1998. Effects of fibrolytic enzyme treatment on forage dry matter and organic disappearance. J. Animal Sci. 76 (Suppl. 1):293 (Abstr.).
TRICARICO, J.M.; DAWSON, K.A.; NEWMAN, K. E. 1998. Effects of an exogenous microbial enzyme preparation (Fibrozyme) on ruminal digestion of fescue hay. J. Animal Sci. 76 (Suppl. 1):289 (Abstr.).
VAN SOEST, P.J.; ROBERTSON, J.B.; LEWIS, B.A. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Anim. Sci. 74:3583-3597.
Juan H. Avellaneda-Cevallos2, Sergio S. González-Muñoz3, Juan M. Pinos-Rodríguez4, Alfonso Hernández-Garay3, Oziel D. Montañez-Valdez5, José Ayala-Oseguera6
2 Unidad de Investigación Científica y Tecnológica, Ftad. de Cs. Pecuarias, Univ. Técnica Estatal de Quevedo. Quevedo, Ecuador.
3 Colegio de Postgraduados. Programa de Ganadería. Montecillo, Texcoco, Estado de México, México. y
4 Instituto de Investigación de Zonas Desérticas, Univ. Autónoma de San Luís Potosí, San Luís Potosí, México.
5 División de Bienestar y Desarrollo Regional. Departamento de Desarrollo Regional. CUSUR. Universidad de Guadalajara. Guzmán, Municipio de Zapotlán El Grande, Jalisco, México. .
6 Departamento de Zootecnia. Universidad Autónoma de Chapingo, Texcoco, Estado de México, México.
2 Unidad de Investigación Científica y Tecnológica, Ftad. de Cs. Pecuarias, Univ. Técnica Estatal de Quevedo. Quevedo, Ecuador.
3 Colegio de Postgraduados. Programa de Ganadería. Montecillo, Texcoco, Estado de México, México. y
4 Instituto de Investigación de Zonas Desérticas, Univ. Autónoma de San Luís Potosí, San Luís Potosí, México.
5 División de Bienestar y Desarrollo Regional. Departamento de Desarrollo Regional. CUSUR. Universidad de Guadalajara. Guzmán, Municipio de Zapotlán El Grande, Jalisco, México. .
6 Departamento de Zootecnia. Universidad Autónoma de Chapingo, Texcoco, Estado de México, México.
Temas relacionados
Autores:

Únete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicación
14 de noviembre de 2007
Me parece interesante el artículo, felicitaciones, sin embargo, valdría la pena argumentar en relación al problema que se suscita en animales que pastan en brizantha, como la fotosensibilización, que trae consigo pérdidas económicas... Es más, se comentó de que era un problema de manejo del pasto.
Espero sus comentarios.



Te puede interesar
