INTRODUCCIÓN
El periodo seco es una de las etapas más críticas en el ciclo productivo de la vaca lechera. Un periodo seco inadecuado puede dañar la producción de leche, la eficiencia reproductiva de la vaca y causar pérdidas económicas al productor. Dicho periodo de estrés incluye eventos como la depresión del consumo de alimento y los cambios homeorréticos y hormonales propiciados por el desarrollo del feto, el parto e inicio de la lactancia (Bauman y Curie 1980). El manejo nutricional para obtener un periodo seco óptimo requiere de un conocimiento amplio de eventos bioquímicos y endocrinos que ocurren en el animal hacia finales de la gestación, las actividades de parto y el inicio de la lactancia.
El manejo nutricional durante el periodo seco tiene un papel fundamental en la reducción del estrés después del parto; el fundamento estriba en suministrar los nutrimentos necesarios para mantener el equilibrio del ambiente uterino que contiene un feto, placenta y líquidos fetales, y para generar reservas corporales que serán removidas rápidamente en la lactancia siguiente. El manejo exitoso deberá minimizar los problemas de salud y maximizar la productividad en la lactancia inmediata. Debido a la asociación entre la acumulación de lípidos hepáticos y desarreglos metabólicos periparturientos, el manejo nutricional debe promover un apetito vigoroso para incrementar el consumo de materia seca (CMS) después del parto. Debido a que el consumo de alimento se deprime en las primeras semanas después del parto y los requerimientos incrementan, el animal moviliza reservas corporales que se asocian con el incremento en la producción de triglicéridos hepáticos (Douglas y col 2007; Litherland y col 2011). En el escenario anterior, la densidad energética de las dietas tiene una función preponderante como regulador del consumo de alimento y promotor de la acumulación de reservas corporales.
La mayoría de los ganaderos lecheros tienen como práctica común limitar el CMS durante el periodo seco para preparar a la vaca para un mayor consumo en la lactancia siguiente; esto es como una estrategía para reducir los trastornos metabólicos. Por otro lado, un CMS alto durante el periodo seco puede incrementar significativamente la acumulación de grasa corporal, la cuál reduce el apetito y minimiza la producción de leche(Rukkwamsuk y col 2000). Ambos aspectos han sido estudiados con cierto detalle; sin embargo, todavía es debatible si la alta (McMillen y col 2001) o baja (Osgerby y col 2002; Quigley y col 2008) densidad de energía preparto tendría efectos negativos en el creciemeinto fetal y la productividad en la lactancia siguiente. Con base en lo anterior, el objetivo del presente estudio fue determinar el efecto de dietas con diversos niveles de ENL en los cambios de peso vivo, condición corporal, consumo de alimento y energía, y la eficiencia de utilización de la energía en vacas Holstein-Friesian durante el periodo seco de 60 días.
MATERIAL Y MÉTODOS
ÁREA DE ESTUDIO
El estudio se realizó en el establo lechero “18 de Julio” propiedad de la Universidad Autónoma Chapingo, ubicado en el Municipio de Tlahualilo, Durango. El sitio se ubica a 25º 54’ 07” N y 103º 35’ 09” O, la altitud es 1,137 m y el clima es desértico. La temperatura media anual es de 21,1°C con una precipitación pluvial de 239 mm anuales, distribuyéndose principalmente de julio a septiembre (García 1988).
ALIMENTACIÓN Y MANEJO DE ANIMALES
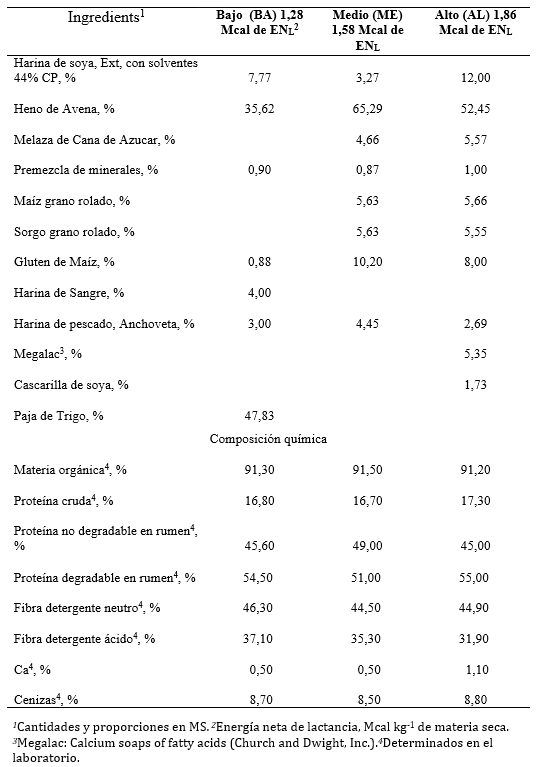
Los animales usados estaban gestantes, de 60 días de la fecha probable de parto. La fecha de inicio del experimento fue fija y sólo se incorporaron tres animales más tarde: 1 un día y 2 tres días después. Las vacas fueron seleccionadas por su estado saludable, y condición corporal 3,15 ± 0,25 (escala de 1, muy delgada; 5, muy gorda). El grupo experimental consistió de 30 animales (n = 10 por tratamiento), de más de dos lactancias con peso vivo (PV) promedio de 667,0 ± 23,3 kg. Posterior al secado, las vacas fueron estratificadas por PV y producción de leche anterior y fueron aleatoriamente asignadas a uno de tres tratamientos con diferentes concentraciones de energía neta de lactancia (ENL). Los tratamientos fueron bajo (BA), medio (ME) y alto (AL) y su contenido fue 1,28, 1,58 y 1,88 Mcal de ENL kg-1 de MS, respectivamente (Cuadro 1). Las dietas fueron elaboradas siguiendo las recomendaciones del NRC (NRC 2001) para vacas en periodo seco. El alimento se ofreció en tres proporciones iguales (33.3% del consumo total) a las 0500, 1300 y 2100 h. Los animales se entrenaron para obtener el alimento de comederos individuales tipo Calan Door™; que consisten de una caja de aproximadamente 36,0 kg donde se depósita el alimento.
Los comederos son exclusivos para cada animal. Las vacas fueron dotados con un medallón que contiene un semiconductor electrónico que coincide con la combinación de la puerta. Una vez que el animal se retira, la puerta se cierra automáticamente y sólo se abrirá cuando el mismo animal intente comer nuevamente. Para facilitar el manejo, las vacas se identificaron con aretes de colores diferentes y con números progresivos del 1 al 30. Las dietas experimentales se suministraron ad libitum durante 60 días antes del parto, y sólo se extendió un par de días más para los animales que entraron más tarde. Los animales tuvieron acceso a agua limpia y fresca a libertad.
CAMBIOS DE PESO Y MEDICIONES
En las primeras horas de la mañana, todos los días, las vacas seleccionaban uno de los 30 comederos, y en función de la vaca y del tratamiento se les asignó el alimento en la cantidad correspondiente. Como se indicó anteriormente, el alimento del días se dividió en tres porciones y cada porción se suministró en el horario designado. El alimento ofrecido fue previamente pesado y registrado; la proporción no consumida se pesó y se registró. Este procedimiento se repetió en cada horario de ofrecimiento. Al final del día se contabilizaron las porciones no consumidas y por diferencia con respecto al ofrecido se determinó el consumo diario individual. El consumo se estableció proporcionando 15,0% más del requerimiento de alimento sugerido por el NRC (NRC 2001). El alimento se ofreció ad libitum durante el día. El alimento no consumido al final de día se removió y se registró como no consumido. Al día siguiente, se le ofreció la cantidad estimada previamente más el 15%, y en forma sucesiva hasta que se estabilizó; lo que ocurrió aproximadamente en seis días.
Las vacas gestantes recibieron tratamiento de secado de ubres 18 días antes del inicio del experimento. Posterior al secado, los animales recibieron una dieta de adaptación con base en heno de alfalfa y maíz rolado durante 14 días. Las vacas se pesaron al inicio del experimento (60 días antes de la fecha probable de parto), cada 14 días durante el experimento y un día antes y después del parto. Dichas mediciones fueron usadas para estimar el PV y la ganancia diaria de peso (GDP) de los animales. Los animales fueron pesados en las primeras horas de la mañana.
El escore de condición corporal (CC) se registró antes del pesaje al inicio del experimento, 30 días después y al final del periodo seco. En todos los casos fue durante las primeras horas del día y antes del ordeño. Para la determinación de la CC se usó una escala de cinco puntos con incrementos intermedios, donde 1 representó a las muy delgadas y 5 a las vacas obesas como lo describió Ferguson y col (1994).
La eficiencia de utilización de la energía se determinó considerando la GDP y el consumo de ENL (von Soosten y col 2012); mientras que el consumo de ENL se determinó multiplicando el CMS por el contenido de ENL de la dieta.
Del alimento ofrecido diariamente a cada vaca se obtuvó una muestra, que al final de la semana se conjuntó y formó una muestra semanal del alimento consumido y del no consumido. En ambas muestras se determinó el contenido de MS y la calidad del alimento durante el periodo experimental. Después de la colección, las muestras de alimento fueron almacenadas a -20 °C, se pesaron y secaron a 55-60 °C en una estufa de aire forzado por 48 h para determinar la MS de campo parcial. Posteriormente, las muestras de alimento se molieron en un molino Willey (A. H. Thomas, Philadelphia, PA, USA). La materia seca total se determinó usando un estufa a 100 °C durante 24 h (AOAC 2000). Por último, las muestras se incineraron en mufla a 500 °C para determinar el contenido de materia orgánica (MO) y cenizas. Los contenidos de fibra detergente neutro (FDN) y ácido (FDA) se determinaron usando los métodos de Goering y Van Soest (1970). Adicionalmente, las muestras se analizaron para determinar el contenido de PC (%) usando el método Kjeldahl (AOAC 2000).
Los ingredientes, la composición química y el valor nutritivo de las dietas experimentales utilizadas se muestran en la Cuadro 1. Los contenidos de MO, PC, FDN, FDA, ENL, minerales y cenizas se determinaron en el laboratorio de acuerdo a los procedimientos descritos anteriormente.
ANÁLISIS ESTADÍSTICOS
Los datos fueron analizados con el procedimiento Mixed de SAS(2010; SAS Institute Inc., Cary, NC; version 9,3). El CMS se determinó diariamente para cada animal, mientras que el PV se registró semanalmente durante el periodo experimental. El PV inicial y el incremento en PV semanal se usaron para estimar los cambios de PV. El CMS, ENL, cambios de PV, CC y la eficiencia de utilización de la energía se analizaron con Mixed de SAS en un diseño completamente al azar de medidas repetidas en el tiempo (SAS 2010). El modelo incluyó los efectos fijos de tratamiento, semana y la interacción tratamiento x semana, con vaca anidada dentro de tratamiento como el término repetido.
El modelo final es como se indica, después de remover las covariables y las interacciones dobles y triples que fueron no significativas (P > 0,05):
Yijkl = μ + TRATi + SEMANAj + TRAT X SEMANAij + Eijkl
Donde: Yijkl es una observación de la variable respuesta; μ,es la media general; TRATi es el efecto fijo del i-ésimo tratamiento (i= 1, 2 y 3); SEMANAj es el efecto fijo de la j-ésima semana de experimentación (1, 2,…8); TRAT X SEMANAij es el efecto fijo de la interacción tratamiento x semana y Eijkl es el error aleatorio.
Un modelo similar al anterior; con diferentes interacciones y covariables se utilizó para analizar las concentraciones de PC, FDA, FDN y MS de los alimentos. De los resultados obtenidos se reportan las medias de mínimos cuadrados y los errores estándar. Los resultados fueron declarados significativos usando una P < 0,05. La opción PDIFF del procedimiento Mixed se utilizó para determinar las diferencias entre tratamientos en puntos específicos (P < 0,05).
Las estructuras de covarianzas consideradas fueron de simetría compuesta y autorregresiva de orden 1. La estructura de covarianza que proporcionó la información más baja de los criterios de Akaike (Litell y col 1996) fue la de simetría compuesta y se usó para cambios de PV, GDP y eficiencia de utilización de la energía. Las variables de consumo de MS y de energía requirieron del uso de la covarianza autorregresiva de orden 1; mientras que la de simetría compuesta fue la más apropiada para PC, FDN, FDA y MS.
RESULTADOS
CAMBIOS DE PESO CORPORAL, METABÓLICO Y CONDICIÓN CORPORAL
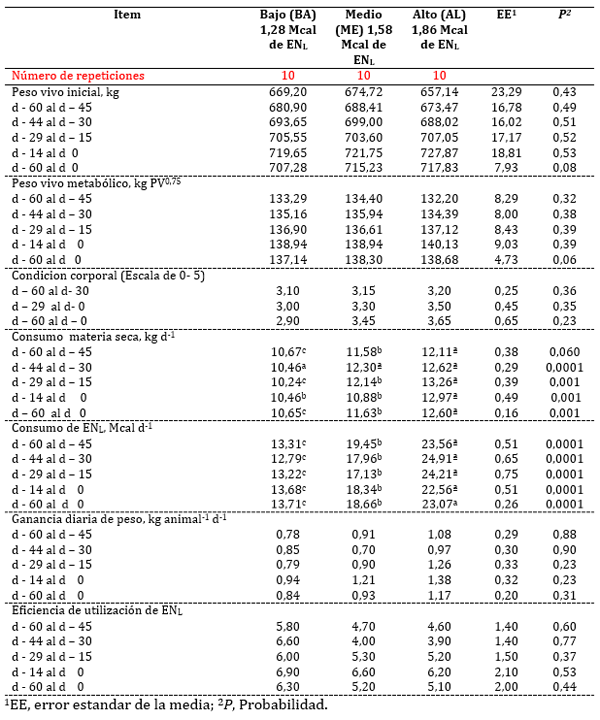
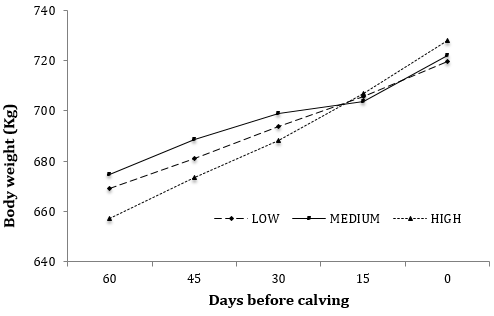
El experimento fue planeado para probar el efecto de diferentes niveles ENL en las dietas ofrecidas a vacas secas en un periodo convencional de 60 días. Para lograr dicho objetivo se estudiaron diferentes variables incluyendo el peso vivo (PV), metabólico (kg de PV0,75) y CC (Cuadro 2). El PV promedio inicial de las vacas fue 667,02 ± 23,29 kg; que como se esperaba no hubo diferencias (P > 0.05) entre grupos experimentales. También se observó que dicha situación se mantuvo sin cambios en la mayoría de los periodos de 14 días analizados. Sin embargo, se observó un efecto del tiempo (P < 0,0001) del d -15 al d - 0 (P < 0,001; Figura 1). Contrariamente, cuando el PV se analizó como un sólo periodo, se observó una tendencia (P = 0,08) de mayor PV para los niveles ME y AL. Sin embargo, las vacas de los niveles AL mostraron incrementos númericos de PV superiores al ME y BA (60,59 vs 41,11 y 38,08 kg, respectivamente) durante el periodo global de 60 días. Las diferencias en PV expresadas en unidades de peso metabólico (kg de PV0,75) también mostraron una tendencia (P = 0,06) de mayor PV en los niveles ME y AL.
No hubo diferencias entre grupos experimentales en CC en el momento del secado (P < 0,36), ni a los 30 dÍas (P < 0,36), tampoco al final del experimento (P < 0,36); sin embargo, al final del periodo experimental, las vacas con el nivel AL mostraron CC mejores comparados con las del ME y BA.
CONSUMO DE MATERIA SECA Y ENERGÍA
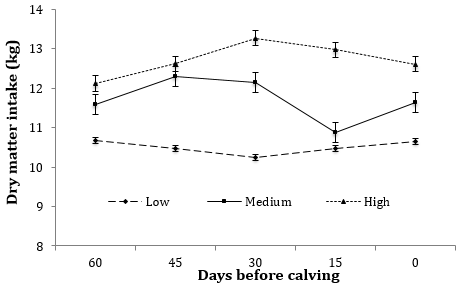
Los consumos de MS y ENL analizados por periodos de 15 días y en forma global durante el periodo seco de 60 días se presentan en el Cuadro 2. En dicho periodo, el efecto de los tratamientos influyó (P < 0,0001) en el CMS y de ENL (P < 0,0001) durante todos los periodos de 15 días, desde el día 60 hasta el 0. El CMS fue 1,436 y 0,537 kg animal-1 d-1 mayor en el nivel AL comparados con ME y BA, respectivamente, del d - 60 al d - 45 (Figura 2). Dicho comportamiento se expresó de manera similar en los periodos siguientes hasta el parto. Con excepción del nivel BA, el CMS disminuyó conforme los animales se acercaban al tiempo de parto.
El consumo de ENL mostró un comportamiento similar al CMS expresado por los animales durante el periodo seco. Las vacas del nivel AL consumen 4,11 y 10,25 más Mcal de ENL kg-1 de alimento comparado con el ME y BA, respectivamente, del día 60 al día-45; dichas diferencias se mantuvieron durante todos los periodos analizados hasta el parto.
GANANCIA DE PESO Y EFICIENCIA DE UTILIZACIÓN DE LA ENERGÍA
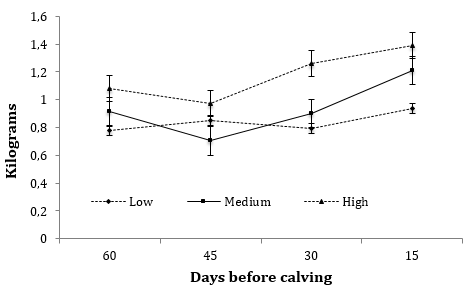
La ganancia diaria de peso (GDP) y la eficiencia de la utilización de la energía por las vacas en periodo seco se presentan en el Cuadro 2. Como se observa en dicho Cuadro, los niveles de energía no influyeron (P > 0.05) en la GDP de los vacas en los distintos periodos de alimentación. Contrariamente, se observó un efecto del tiempo (P < 0,0001) y de la interacción tratamiento x tiempo (P < 0,001) en el periodo del d - 60 al d - 45. Las vacas del nivel AL fueron 9,9 y 20,0% más pesadas comparadas con los tratamiento ME y BA, respectivamente (Figura 3).
La eficiencia de utilización de la energía (GDP/Mcal de ENL consumida) mostró un comportamiento distinto en GDP. Los diferentes niveles de ENL incluídos en las dietas no influyó (P = 0,44) en la eficiencia de utilización en los distintos grupos experimentales. Contrariamente, en todos los periodos de 15 días se encontró un efecto significativo (P < 0,001) del tiempo; mientras que el periodo del d - 44 al d - 30 se encontró una interacción tratamiento x tiempo significativa (P < 0,02). Las vacas del nivel AL fueron 19,2 y 17,5% más eficientes en la utilización de la ENL comparadas con las del ME y BA, respectivamente (Figura 4).
DISCUSIÓN
CAMBIOS DE PESO CORPORAL Y METABÓLICO Y CONDICIÓN CORPORAL
Una de las preocupaciones básicas de los productores lecheros es encontrar el manejo apropiado para optimizar los nutrimentos durante el periodo seco, sin perjudicar la salud de los animales y facilitar la transición a la lactancia posterior. En dicho contexto, una de las interrogantes más frecuentes es el nivel de ENL que debe proveerse a la vaca; lo que condujo a la formulación de dietas con diferentes niveles de energía durante un periodo seco de ocho semanas.
Los niveles diversos de energía preparto no generaron diferencias en cambios de PV, PV0.75 y CC durante el periodo seco. El incremento en PV de 38,08 y 40,51 kg de los grupos BA y ME, respectivamente, probablemente, represente una disminución en la masa corporal, considerando que el peso de la placenta, el feto y los fluídos fetales incrementan aproximadamente 45,0 kg durante las ocho semanas (Klop y col 1998). Como consecuencia, las vacas del nivel BA perdieron 0,20, mientras que las del ME y AL incrementaron en 0,20 y 0,40, respectivamente, unidades de CC.
La mayor concentración de energía de los niveles BA y ME fue usada, posiblemente, para formación del feto, que aunque no hubo diferencias estadisticas en el PV de los terneros al nacimiento (45,60, 47,23, y 50,45 ± 8.40 kg, para BA, ME y AL, respectivamente; P > 0,05), los del ME y AL fueron más pesados. También se observó que los PV postparto fueron similares (645,30, 670,70 y 659,80 ± 20,2 kg; para BA, ME y AL, respectivamente). Una proporción menor de energía, posiblemente, se usó para la regeneración de tejido adiposo; que aunque los análisis estadísticos utilizados no fueron significativos(3,20, 3,45 y 3,65 ± 0,33, para ME, BA y AL, respectivamente; P = 0,23; Cuadro 2), la observación visual de las vacas del ME y sobre todo las del AL se observaron con mejores reservas corporales.
Agënas y col (2003) observaron que las vacas que recibieron concentraciones de ENL (13,56, 20,25 y 33,82 Mcal d-1, niveles BA, ME y AL, respectivamente) similares a las del presente estudio
(13,63, 18,37 y 23,43 Mcal d-1 para los mismos niveles) obtuvieron resultados análogos en cambios de PV y CC. Resultados contradictorios fueron observados por Dann y col (2006), quienes indicaron que las vacas que recibieron dietas completas con 1,30 y 1,59 Mcal kg-1 de ENL promovieron cambios en el PV de 2,0, y 28,0 kg, respectivamente, del dia 60 al d-25 del periodo seco. Los incrementos en el PV, no son consistentes con los observados en el presente estudio.
El escore de CC es una herramienta que puede usarse en los hatos modernos donde el volumen de animales no permite tener un seguimiento puntual de cada vaca. Sin embargo, la subjetividad de la evaluación incrementa la variabilidad y genera dudas de la confiabilidad del escore. Por otro lado, también existe la necesidad de mejorar los conocimientos de movilización de reservas corporales. En el presente estudio, las vacas iniciaron el secado con similar CC y a pesar de que hubo incrementos en los niveles ME y AL durante el periodo seco, es posible, que las vacas del ME hayan presentado movimientos de grasa interna sin que fuera detectado; posiblemente, por el periodo relativamente corto de estudio. Otro factor de confusión, eventualmente, lo represente las diferencias en la talla corporal de los animales, debido a que los PV no fueron diferentes; sin embargo, los animales de talla más grande consumieron más alimento. Pires y col (2013) observaron que las vacas que consumieron dietas de 1,40 Mcal del secado hasta la semana 3 y posteriormente de 1,45 Mcal de energia neta hasta el parto, y con CC de 3,0, 3,2-3,8 y > 4,0, para bajo, medio y alto, respectivamente, mostraron comportamientos en PV y consumos de alimento similares a los observados en el presente estudio.
CONSUMO DE MATERIA SECA Y ENERGÍA
El incremento en la densidad energética de las dietas durante las últimas ocho semanas de periodo seco aumentó el CMS y energía(Cuadro 2); lo que concuerda con otros resultados publicados anteriormente (Keady y col 2001; Agënas y col 2003; Rabelo y col 2003). Las vacas del nivel AL consumen 8,44 y 16,00% más MS y 27,0 y 41,0% más energía que las del ME y BA, respectivamente. Dicha diferencia se explica parcialmente por el contenido de fibra; las dietas del nivel AL (45,00 vs 49,00 y 46.30 de fibra detergente neutro, respectivamente) fueron más bajas en fibra comparadas con las del ME y BA, lo que permitió una digestión ruminal más alta, seguido por una evacuación más rápida de la digesta. Rabelo y col (2003) reportaron consumos de alimento de 11,3 y 13,0 kg d-1 y de energía de 17,7 a 21,5 Mcal en vacas preparto. Las vacas recibieron dietas con 1,55 Mcal del día 35 hasta el 14 antes del parto. Posteriormente, recibieron dietas con 1,65 Mcal del día 14 hasta el parto. Los autores concluyeron que incrementando la densidad de la energía en las dietas preparto se logró un incremento en el consumo de alimento y energía.
El consumo extra de alimento y energía relativo a los requerimientos demostró que las vacas secas y preñadas no regulan el CMS y de energía, a menos de que se preparen para ganar CC. En el presente estudio, las vacas del AL consumieron un promedio de 2,0% de su PV como MS, confirmando otros reportes de que las vacas secas tienen la habilidad de consumir alimento en proporciones grandes aún con contenidos altos de energía (Grummer y col 2004; Douglas y col 2006).
VandeHaar y col (1999) indicaron que el incremento en la densidad de energía de 1,3 a 1,6 Mcal de ENL kg-1 incremento el consumo de alimento en 14% y de energía en 40% durante las dos últimas semanas de gestación. Lo que concuerda con los resultados obtenidos en el presente estudio. VandeHaar y col (1999) también indicaron que el consumo de alimento de animales con bajos requerimientos de nutrimentos frecuentemente decrecen cuando la concentracion de energía se incrementa. En el presente estudio, se observó un efecto distinto; la mayor concentración de energía permitió consumos de 1,0 y 2,0 kg animal-1 d-1 más para ME y BA, respectivamente. Una diferencia, posiblemente, se deba a que el estudio de VandeHaar y col (1999) sólo se consideró las dos últimas semanas antes de parto, mientras que el presente estudio se consideró un tiempo de ocho semanas.
Algunos reportes de VandeHaar y col(1999), Grummer y col(2004) y Janovich y col(2010) indican que el CMS de vacas gestantes tiende a reducirse conforme los animales se acercan al parto. La disminución puede ser ligera (Grum y col 1996; aproximadamente el 10% de reducción) o más severa (Grummer y col 2004; Douglas y col 2006; aproximadamente el 30 y 47% de reducción). En el presente estudio, el consumo de energía también se redujó en las dos últimas semanas de gestación. Dicho efecto fue más visible en los niveles ME yAL, que aunque no fue significativo, mostró una tendencia a la reducción en las dos últimas semanas, en concordancia con los reportes de la literatura mencionados en párrafos anteriores.
También fue de interés del estudio determinar los cambios en CC cuando las vacas tienen restricciones o excesos de alimento durante el periodo seco, de acuerdo con las recomendaciones del NRC (NRC, 2001). Las dietas usadas se formularon para vacas de 650 kg de PV, con ganancias de aproximadamente 36 kg por arriba del crecimiento del feto, o 0,6 unidades de CC. El plan inicial consideró que las vacas del BA perderían, que las del ME mantendrían y las del AL ganarían CC. Los resultados demuestran que las vacas del BA efectivamente perdieron CC; mientras que las del ME y AL ganaron 0,20 y 0,45 unidades, respectivamente, hasta que alcanzaron el parto (Holcomb y col 2001). Los resultados del presente estudio son contrarios a los observados por Grum y col (1996). En dicho estudio, las dietas contenían forrajes de calidad y digestibilidad diferente a los utilizados en el presente estudio, lo pudo haber motivado una respuesta diferente.
GANANCIA DE PESO Y EFICIENCIA DE UTILIZACIÓN DE ENERGÍA
La densidad energética de las dietas no influyó en la GDP de los animales durante el periodo experimental, lo que concuerda con los resultados observados por Douglas y col (2006). La deferencia númerica se explicó por el consumo de alimento mostrado por las vacas. Dicho consumo proprcionó, en promedio, un diferencial de energía de aproximadamente 4,41, 4,95 y 9,36 Mcal del AL al ME, y de este último al BA y del AL al BA, respectivamente; esto se explica, posiblemente, porqué el suministro de las dietas fue en un periodo relativamente corto. También se observó que los cambios de PV sólo mostraron una tendencia (P = 0,08) de superioridad de los niveles AL y ME (60,69 y 40,51 vs 38,08, ± 8,40 kg, respectivamente) con respecto al BA. Los resultados obtenidos en el presente estudio son similares a los observados por Agënas y col (2003), aunque en dicho estudio, los animales iniciaron con CC más altas 10 semenas antes del parto.
Debido a que los PV de los terneros al nacimiento no fueron diferentes (45,60, 47,23, y 50,45 ± 8,58 kg, para BA, ME y AL, respectivamente; P = 0,40), y asumiendo que el peso de los tejidos y fluídos fetales incrementa aproximadamente 45,0 kg durante un periodo de ocho semanas(Law y col 2011); las vacas de los niveles BA y ME, posiblemente, movilizaron más tejido materno comparadas con el AL, para apoyar el desarrollo fetal durante el periodo seco.
En el presente estudio, la variacion en el contenido de ENL de las dietas durante el periodo seco no infuyó la eficiencia de utilización de la energía (Cuadro 2). La diferencia en la concentración de energía del BA al AL fue de 0,58 Mcal kg-1 d-1 aproximadamente; lo que concuerda con el uso de la energía y su relación con el consumo de alimento expuesto en líneas anteriores (Gao y col 2009; Micke y col 2010).
Las vacas en los grupos BA, ME y AL presentaron tamaños metabólicos corporales similares en el periodo seco (137,14, 137,15 y 138,68 ± kg de PV0,75, respectivamente); lo que sugiere un efecto de la diferencia en energía. Aparentemente, el nivel BA (1,28 Mcal) promovió mayores formaciones de tejido fetal a costa de la remoción de reservas corporales de la vaca preparto (Robinson y col 1999; McMillen y col 2001) mientras que los niveles ME y AL destinan más energía para formación de reservas corporales.
Aunque los PV de las vacas fueron similares durante el perido seco, se esperaría que las del nivel AL parieran terneros más pesados que los demás niveles. Gao y col (2012) indicaron que densidades de energía de 1,28 Mcal de ENL (similar al nivel BA de presente estudio) no afectaron el PV de las vacas en los últimos 21 de gestación; sin embargo, las concentraciones de metabólitos sanguíneos como glucosa, NEFA y BHBA se alteraron, lo que cambió la relación de la vaca con el feto y promovió una reducción en el peso del mismo; dicho efecto no fue evidente en el presente estudio. Los mismos autores indicaron que la dieta con energía alta (1,58 Mcal; que coincide con el nivel ME utilizado en el presente estudio), mejoró el crecimiento postnatal y la salud de los terneros.
Los resultados del presente estudio requieren observarse con precaución debido al número pequeño de vacas utilizadas por tratamiento (10 animales) y debido a que no determinaron metabólitos y hormonas relacionados con la movilización de reservas y utilización del alimento.
A pesar de lo anterior, el presente estudio propone algunas consideraciones de manejo. La primera se relaciona con las dietas de 1,28 Mcal de ENL por kg de alimento fueron suficientes para mantener GDP superiores a 0,8 kg animal-1 día-1 y producir terneros de aproximadamente 45,0 kg al nacimiento en vacas con periodos secos de 60 días y que al secado tenían 3,10 de CC. Otra consideración, se relaciona con un nivel de 1,86 Mcal en la dieta y vacas con CC de 3,10 al secado, no solamente mantienen GDP superiores que 1,2 kg animal d-1 sino también acumulan aproximadamente 70,73 ± 7,9 kg por animal-1 d-1 de reservas corporales que serán removidas al inicio de la lactancia, sin la presencia de trastornos metabólicos aparentes. El exceso de energía no tuvo efectos negativos en la presentación del celo de las vacas (41,60, 54,70 y 70,00 ± 34,30 días para BA, ME y AL; respectivamente; P > 0,05) ni en la capacidad para preñarse nuevamente (días abiertos, 106,75, 164,90 y 143,44 ± 33,48 para BA, ME y AL, respectivamente; P > 0,05), como fue sugerido por Rukkwamsuk y col(2000) para animales con ganancias y condiciones similares a las del presente estudio.
CONCLUSIONES
Las dietas con 1,28 Mcal de energía neta de lactancia y condición corporal 3,10 al secado son apropiadas para vacas durante el periodo seco, mantienen el peso corporal de los animales, la gestación, y el crecimiento del feto.
Las dietas con 1,86 Mcal de energía neta de lactancia y condición corporal 3,20 al secado incrementan las reservas corporales y el crecimiento del feto y no compremeten la eficiencia reproductiva posparto.
REFERENCIAS
Agënas S, E Burstedt, K Holtenius. 2003. Effects of feeding intensity during the dry period. 1. Feed intake, body weight, and milk production. J Dairy Sci 86, 870-882.
AOAC. 2000. Official Methods of Analysis. Association of Official Analytical Chemists. 17th Edition. Association of Official Analytical Chemists Press. Gaithersburgh, MD.
Bauman DE, WB Currie. 1980. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J Dairy Sci 63, 1514-1529.
Dann HM, NB Litherland, JP Underwood, M Bionaz, A D’Angelo, JW McFadden, KJ Drackley. 2006. Diets during far-off and close-up dry periods affect parturient metabolism and lactation in multiparous cows. J Dairy Sci 89, 3563-3577.
Dann HM, MP Carter, KW Cotanch, CS Ballard, T Takano, RJ Grant. 2007. Effect of partial replacement of forage neutral detergent fiber with by-product neutral fiber detergent fiber in close-up diets on periparturient performance of dairy cows. J Dairy Sci 90,1789-1801.
Douglas GN, TR Overton, HG Bateman II, HM Dann, JK Drackey. 2006. Prepartal plane of nutrition, regardless of dietary energy source, affects parturient metabolism and dry matter intake in Holstein cows. J Dairy Sci 89, 2141-2157.
Douglas GN, J Rehage, AD Beulieu, OA Bahaa, JK Drackley. 2007. Prepartum nutrition alters fatty acid composition in plasma adipose tissue, and liver lipids of periparturient dairy cows. J Dairy Sci 90, 2941-2959.
Ferguson JD, DT Galligan, N Thomsen. 1994. Principal descriptors of body condition score in Holstein cows. J Dairy Sci 77, 2695-2703.
Gao F, YC Liu, ZH Zhang, CZ Zhang, HW Su, SL Li. 2012. Effect of prepartum maternal energy density on growth performance, immunity, and antioxidation capability of neonatal calves. J Dairy Sci 95, 4510-4518.
Gao F, YC Liu, HZ Hou. 2009. Effect of maternal undernutrition during late pregnancy on growth and development of ovine fetal visceral organs. Asian-australasianJ Anim Sci 22, 1633-1639.
García E. 1988. Modificaciones al Sistema de Clasificación Climática de Köppen. UNAM Press. Instituto de Geografía, Universidad Nacional Autónoma de México. México, D. F. 108. p.
Goering HK, PJ Van Soest. 1970. Forage Fiber Analyses (Apparatus, reagents, procedures, and some applications). Agric. Handbook No. 379. ARS-USDA. ARS-USDA Press Inc Washington, DC. (USA).
Grummer RR, DG Mashek, A Hayirly. 2004. Dry matter intake and energy balance in the transition period. Vet Clin North Am Food Anim Pract 20, 447-470.
Grum DE, JK Drackley, RS Younker, DW LaCount, JJ Venhuizen. 1996. Nutrition during the dry period and hepatic lipid metabolism of peripartum dairy cows. JDairy Sci 79, 1850-1864.
Guo J, RR Peters, RA Kohn. 2007. Effect of a transition diet on production performance and metabolism in periparturient dairy cows. J Diary Sci 90, 5247-5258.
Holcomb CS, HH Van Horn, HH Head, MB Hall, CJ Wilcox. 2001. Effects of prepartum dry matter intake and forage percentage on potpartum performance of lactating dairy cows. J Dairy Sci 84, 2051-2058.
Janovich NA, JK Drackley. 2010. Postpartum dietary management of energy intake and lactation performance by primiparous and multiparous Holstein cows. J Dairy Sci 93, 3086-3102.
Keady TW, CS Mayne, DA Fitzpatrick, MA McCoy. 2001. Effect of concentrate feed level in late gestation and subsequent milk yield, milk composition, and fertility of dairy cows. J Dairy Sci 84, 1468-1479.
Klop A, GAL Meijer, TAM Kruip. 1998. Energy intake during the dry period and energy balance in dairy cows around calving. 10th Int. Conf. Prod Diseases Farm Anim. 1995. Abstr.
Law RA, FJ Young, DC Patterson, DJ Kilpatrick, ARG Wylie, KL Ingvarsten, A Hamelleers, MA McCoy, CS Mayne, CP Ferris. 2011. Effect of precalving and postcalving energy level on performance and blood metabolite concentration of dairy cows throughout lactation. J Dairy Sci 94, 808-823.
Littell CR, AG Milliken, WW Stroup, FD Wolfinger. 1996. SAS System for Mixed Models. SAS Inst., Inc. Press. Cary, (North Carolina) USA.
Litherland WB, HM Dann, JK Drackley. 2011. Prepartum nutrient intake alters palmitate metabolism by liver slices from peripartal dairy cows. J Dairy Sci 94, 1928-1940.
McMillen IC, MB Adams, JT Ross, CL Coulter, G Simoneta, JA Owens, JS Robinson, LJ Edwards. 2001. Fetal growth restriction: Adaptations and consecuences. Reproduction 122, 195-204.
Micke GC, TM Sullivan, RJ Soares Magalhaes, PJ Rolls, ST Norma, VEA Perry. 2010. Heifer nutrition during early- and mid-pregnancy alter fetal growth trajectory and birth weight. Anim Rep Sci 117, 1-10.
NRC. 2001. National Research Council. Nutrients requirements of dairy cattle. 7th ed. National Academic Press. Washington, D. C. 381 pp.
Osgerby JC, DC Wathes, D Howard, TS Gadd. 2002. The effect of maternal undernutrition on ovine fetal growth. J Endocrinol 173, 131-141.
Pires JAA, C Delavaud, Y Faulconnier, D Pomiés, Y Chilliard. 2013. Effect of body condition score at calving on inidicators of fat and protein mobilization of periparturient Holstein-Friesian cows. J Dairy Sci 96, 6423-6439.
Quigley SP, DO Kleeman, SK Walker, PA Speck, SR Rudiger, GS Nattrass, MJ De Blasio, JA Owens. 2008. Effect of variable long-term maternal fetal allowance on the development of the ovine placenta and fetus. Placenta 29, 539-548.
Rabelo E, RL Rezende, SJ Bertics, RR Grummer. 2003. Effects of transition diets varying in dietary energy density on lactation performance and ruminal parameters of dairy cows. J Dairy Sci 86, 916-925.
Robinson JJ, KD Sinclair, TG MCEvoy. 1999. Nutritional effect on foetal growth. Anim Sci 68, 315-331.
Rukkwamsuk T, MJH Geelen, TA Kruip, T Wensing. 2000. Interrelation of fatty acid composition in adipose tissue, serum and liver of dairy cows during the development of fatty liver pospartum. J Dairy Sci 83, 52-59.
SAS. 2012. SAS User’s Guide: Statistics (Version 9.1.3). SAS Inst. Inc. Cary, NC, USA.
VandeHaar MJ, G Yousif, BK Sharma, TH Herdt, RS Emery, MS Allen, JS Liesman. 1999. Effect of energy and protein density of prepartum diets on fat and protein metabolism of dairy cattle in the parturient period. J Dairy Sci 82, 1282-1295.
von Soosten D, V Meyer, M Piechotta, G Flachowsky, S Dänicke. 2012. Effects of conjugated linoleic acid supplementation on body compoition, body mobilization, protein accretion, and energy utilization in early lactation dairy cows. J Dairy Sci 95,1222-1239.
Cuadro 1. Ingredientes, composición química y valor nutritivo de las dietas completas utilizadas por vacas Holstein-Friesian durante un periodo seco de 60 días en confinamiento
Cuadro 2. Medias de minimos cuadrados, error estandar (EE) y nivel de significancia de (P) de peso corporal, ganancia diaria de peso, consumo y eficiencia de utilizacion de ENL de vacas Holstein-Friesian que consumen diferentes niveles de energia durante 60 d antes del parto
Figura 1. Medias de minímos cuadrados de Pesos Vivos catorcenales (Kg) de vacas Holstein-Friesian que recibieron diferentes niveles ENL durante 60 días de periodo seco en confinamiento. Los niveles nutricionales fueron: Bajo, Medio y Alto que consisten en 128, 1,58 y 1,86 Mcal de ENL kg-1 de alimento, respectivamente. El EEM varió de 7,93 a 23,29. Efectos en el modelo: Nivel de energía: P < 0,08; semana, P < 0,57; tratamiento x semana, P < 0.08
Figura 2. Medias de minímos cuadrados de Consumo de materia seca catorcenales (kg) de vacas Holstein-Friesian que recibieron diferentes niveles ENL durante 60 días de periodo seco en confinamiento. Los niveles nutricionales fueron: Bajo, Medio y Alto que consisten en 1,28, 1,58 y 1,86 Mcal de ENL kg-1 de alimento, respectivamente. El EEM varió de 0,16 a 0,38, Efectos en el modelo: Nivel de energía: P < 0,0001; semana, P < 0,028; tratamiento x semana, P < 0,05.
Figura 3. Medias de minímos cuadrados de ganancia diaria de peso catorcenales (Kg) de vacas Holstein-Friesian que recibieron diferentes niveles ENL durante 60 días de periodo seco en confinamiento. Los niveles nutricionales fueron: Bajo, Medio y Alto que consisten en 1,28, 1,58 y 1,86 Mcal de ENL kg-1 de alimento, respectivamente. El EE varió de 0,23 a 0,89. Efectos en el modelo: Nivel de energía: P = 0,31; semana, P < 0,03; tratamiento x semana, P < 0,03.