Potencial de la monensina para reducir las emisiones de metano por parte de la ganadería
Publicado: 30 de diciembre de 2005
Por: Ing. Agr. Darío Colombatto, PhD. Depto. de Producción Animal, Facultad de Agronomía, Universidad de Buenos Aires
Existe un creciente interés en reducir el potencial de calentamiento global, causado por el incremento de la temperatura atmosférica, a través de la reducción de las emisiones de gases con efecto invernadero (GEI). Por ejemplo, el Protocolo de Kyoto establece en sus considerandos que las emisiones de GEI deben reducirse en un promedio de 5.2% por debajo de los niveles de 1990 durante el período 2008-2012 para los países industrializados (Moss, 2002). Los principales gases que integran la categoría de GEI son el dióxido de carbono (CO2), el metano (CH4), el óxido nitroso (N2O), los llamados clorofluorcarbonados (CFC-11 y CFC-12) (Moss et al., 2000). Estos gases difieren en sus características generales y tendencias en función del tiempo (Tabla 1). En el presente trabajo nos concentraremos solamente en el gas metano.
El metano es un potente GEI, ya que su potencial de absorción de radiación es aproximadamente 21 veces superior al del CO2 (Moss et al., 2000) (Tabla 1). Esto trae como consecuencia que, si bien su concentración con respecto al CO2 es muy baja, su contribución al efecto invernadero es importante. Asimismo, dado que su tiempo medio de vida atmosférica es corto, disminuciones en su concentración pueden ser notadas en corto tiempo.
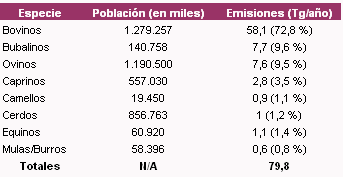
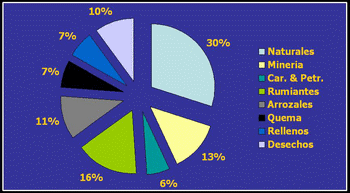
Las fuentes de emisión de gas metano se pueden separar en una primera instancia en antropogénicas o no antropogénicas, contribuyendo las primeras en un 70% del total de las emisiones (Johnson y Johnson, 1995; Kurihara y Terada, 2001). Dentro de las emisiones globales de metano, que totalizan aproximadamente 535 Tg/año (g x 1012), alrededor del 16% está explicado por la fermentación entérica de los animales (Figura 1) (Crutzen et al., 1986; Leng, 1993), contribuyendo los rumiantes (bovinos, bubalinos, ovinos, caprinos en un 95% (US EPA, 1994) (Tabla 2). Algunos países como Australia, Nueva Zelanda, Irlanda, Inglaterra y Canadá realizan esfuerzos para cuantificar el impacto de la fermentación entérica en las emisiones de metano (Hegarty, 2002; Moss, 2002; McCrabb, 2002; O´Hara et al., 2003; Beauchemin and McGinn, 2005). La Argentina, en tanto, ha estimado sus valores de emisiones de metano, basadas primordialmente en modelos estadísticos y ecuaciones matemáticas, siendo las mismas de 2.44 Tg/año (US EPA, 1994).
Tabla 1. Concentraciones, vida media atmosférica y tendencias de los principales GEI, (IPCC, 1990)
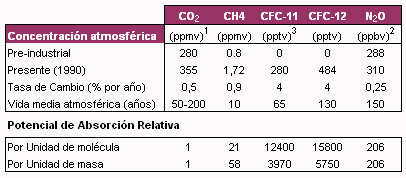
1 Partes por millón en volumen, 2 Partes por billón en volumen, 3 Partes por trillón en volumen
Tabla 2. Poblaciones globales de especies animales domésticas y emisiones de metano en 1990 (US EPA, 1994)
El metano es un potente GEI, ya que su potencial de absorción de radiación es aproximadamente 21 veces superior al del CO2 (Moss et al., 2000) (Tabla 1). Esto trae como consecuencia que, si bien su concentración con respecto al CO2 es muy baja, su contribución al efecto invernadero es importante. Asimismo, dado que su tiempo medio de vida atmosférica es corto, disminuciones en su concentración pueden ser notadas en corto tiempo.
Las fuentes de emisión de gas metano se pueden separar en una primera instancia en antropogénicas o no antropogénicas, contribuyendo las primeras en un 70% del total de las emisiones (Johnson y Johnson, 1995; Kurihara y Terada, 2001). Dentro de las emisiones globales de metano, que totalizan aproximadamente 535 Tg/año (g x 1012), alrededor del 16% está explicado por la fermentación entérica de los animales (Figura 1) (Crutzen et al., 1986; Leng, 1993), contribuyendo los rumiantes (bovinos, bubalinos, ovinos, caprinos en un 95% (US EPA, 1994) (Tabla 2). Algunos países como Australia, Nueva Zelanda, Irlanda, Inglaterra y Canadá realizan esfuerzos para cuantificar el impacto de la fermentación entérica en las emisiones de metano (Hegarty, 2002; Moss, 2002; McCrabb, 2002; O´Hara et al., 2003; Beauchemin and McGinn, 2005). La Argentina, en tanto, ha estimado sus valores de emisiones de metano, basadas primordialmente en modelos estadísticos y ecuaciones matemáticas, siendo las mismas de 2.44 Tg/año (US EPA, 1994).
Tabla 1. Concentraciones, vida media atmosférica y tendencias de los principales GEI, (IPCC, 1990)
1 Partes por millón en volumen, 2 Partes por billón en volumen, 3 Partes por trillón en volumen
Tabla 2. Poblaciones globales de especies animales domésticas y emisiones de metano en 1990 (US EPA, 1994)
Producción de Metano en el Rumen
La fermentación microbiana que ocurre en rumen transforma los carbohidratos, proteínas y glicerol a acetato, dióxido de carbono y amoníaco, produciéndose además metano, propionato y butirato como resultado de reacciones de transferencia de protones y electrones (Van Nevel y Demeyer, 1995). El metano es un producto de desecho nutricional (O´Kelly y Spiers, 1992; Lana et al., 1998), y puede representar entre el 2 y 12% de la energía bruta consumida por el rumiante (Johnson y Johnson, 1995). La reacción química que lleva a la producción de metano es la siguiente (Van Soest, 1994):
4 H2 + CO2 → CH4 + 2 H2O (1)
Excelentes revisiones bibliográficas sobre los aspectos dietarios, ambientales y microbiológicos de la producción de metano en rumiantes han sido publicadas (Johnson y
Johnson, 1995; McAllister et al., 1996).
Figura 1. Contribución de la fermentación entérica a las emisiones de metano (Kurihara y Terada, 2001)
La fermentación microbiana que ocurre en rumen transforma los carbohidratos, proteínas y glicerol a acetato, dióxido de carbono y amoníaco, produciéndose además metano, propionato y butirato como resultado de reacciones de transferencia de protones y electrones (Van Nevel y Demeyer, 1995). El metano es un producto de desecho nutricional (O´Kelly y Spiers, 1992; Lana et al., 1998), y puede representar entre el 2 y 12% de la energía bruta consumida por el rumiante (Johnson y Johnson, 1995). La reacción química que lleva a la producción de metano es la siguiente (Van Soest, 1994):
4 H2 + CO2 → CH4 + 2 H2O (1)
Excelentes revisiones bibliográficas sobre los aspectos dietarios, ambientales y microbiológicos de la producción de metano en rumiantes han sido publicadas (Johnson y
Johnson, 1995; McAllister et al., 1996).
Figura 1. Contribución de la fermentación entérica a las emisiones de metano (Kurihara y Terada, 2001)
Sin embargo, la producción de metano cumple un objetivo importante en el rumen, ya que ayuda a mantener la presión parcial de hidrógeno en un nivel muy bajo, lo que evita que lactato o etanol sean los mayores productos de fermentación, permitiendo que se forme más acetato (Wolin y Miller, 1988). Esto conlleva a una mayor eficiencia del crecimiento bacteriano (Van Nevel y Demeyer, 1995) como resultado de la formación de metano en el rumen. En adición a eso, todas las reacciones del metabolismo ruminal son partes integradas en una serie de reacciones interdependientes entre sí. Por ejemplo, la maximización de la degradación de la fibra está asociada con una mayor producción de metano (Dong et al, 1999). Siguiendo esta línea de razonamiento, uno podría concluir que la eliminación de la producción de metano en el rumen no puede ser considerada una intervención separada del resto, y que sus consecuencias sobre el metabolismo animal y la fermentación del rumen en particular no pueden ser ignoradas (Van Nevel y Demeyer, 1995).
A pesar de lo mencionado, los aspectos negativos de la producción de metano en rumen (pérdidas de eficiencia en el uso de la energía por parte de los animales y contribución al efecto invernadero) han motivado un creciente interés en investigar posibles formas de disminuir las emisiones. Desde el punto de vista de la adopción de tecnologías por parte del productor de carne o leche, es claro que un enfoque que se base en las pérdidas de energía dietaria por parte de los animales, que influyen directamente sobre los márgenes económicos que obtiene cada productor, tendrá un efecto mucho más “convincente” que el enfoque de la disminución de la emisión de GEI para evitar el calentamiento global. Czerkawski (1986) concluyó que las pérdidas de eficiencia en el uso de la energía provocadas por las emisiones de metano significaba una pérdida anual de 15.000 millones de libras esterlinas. Cuantificar la contribución de las emisiones de metano por parte de la ganadería al calentamiento global aparece como más dificultoso (Van Nevel y Demeyer, 1995). Sin embargo, un enfoque integrado debería ser tomado en cuenta por gobiernos o formadores de opinión para desarrollar políticas que contribuyan a un incremento en la productividad ganadera en una forma sostenible en el tiempo.
Estrategias para mitigar las emisiones de metano
Está claro que el primer paso para desarrollar estrategias para disminuir las emisiones de metano debería ser lograr estimadores confiables para cuantificar las emisiones por parte de los animales. Al respecto, existen variados métodos, individuales o grupales, que escapan al objetivo de este trabajo pero que pueden ser consultados por el lector interesado (Johnson et al., 1994; Johnson y Johnson, 1995; McCaughey et al., 1997; McGinn et al., 2004). Las guías del Panel Intergubernamental del cambio climático
(IPCC según sus siglas en inglés) contiene una serie de ecuaciones para la cuantificación de las emisiones de metano por parte de la ganadería (IPCC, 2000). En su versión más simplificada, el método IPCC se basa en definir un factor de emisión de metano para una población específica de animales, y multiplicar este factor por el total de animales que componen la población bajo estudio. Una versión un poco más compleja requiere mayor cantidad de datos de requerimientos, consumo y tasa de conversión de metano (expresadas en unidades de energía por cada 100 unidades de energía bruta consumida) de tipos específicos de alimentos.
i) Intervenciones dietarias
Investigaciones de los años ´60 demostraron que la producción de metano
(expresado como porcentaje de la energía del alimento) estaba inversamente relacionada con el nivel de consumo voluntario de alimentos (Blaxter y Clapperton, 1965). Este concepto se ilustra claramente en la Tabla 3, en donde se puede ver que animales consumiendo dietas basadas en silaje de maíz o ryegrass anual, a niveles de mantenimiento o a 3 veces el nivel de mantenimiento, disminuían sus emisiones de metano en un 39 y 22% con silaje de maíz y ryegrass anual, respectivamente (Kurihara y Terada, 2001). Johnson y Johnson (1995) concluyeron que el porcentaje de energía bruta del alimento perdido como metano disminuía 1.6 unidades porcentuales por cada unidad de incremento del consumo por encima del nivel de mantenimiento. Asimismo, la tasa de fermentación de los sustratos es importante: aquellos alimentos que se degradan a tasas más lentas (caso de los carbohidratos estructurales como la celulosa), producen más metano que aquellos que se fermentan a tasas más rápidas (McCrabb, 2002; Eun et al., 2004).
En general, la inclusión de concentrados en dietas de rumiantes trae aparejado una disminución en el pH ruminal, lo que redunda en una disminución de la relación acético:propiónico y de la producción de metano (Van Kessel y Russell, 1996; Lana et al., 1998; Russell, 1998). En el rumen, las bacterias usan metanogénesis o producción de propionato como estrategias para desligarse de poder reductor, siendo la relación entre ambos (metano y propionato) inversa (Lana et al., 1998).
En función de lo expuesto, Van Nevel y Demeyer (1995) concluyeron que toda aquella intervención dietaria (cantidad/calidad) que produjera una alteración en los perfiles de ácidos grasos volátiles producidos a nivel ruminal (más propionato y menos acetato) impactaría sobre la producción de metano. Al respecto, ejemplos de intervenciones incluyen:
• Incrementar el nivel de carbohidratos de alta fermentabilidad en la dieta, en desmedro de forrajes groseros (McCrabb et al., 1998; Eun et al., 2004).
• Tratamientos físicos de forrajes (molido, peleteado) o de granos (tratamientos con vapor) (Zinn, 1987).
• Tratamientos con urea y minerales en dietas de baja calidad (Leng, 1993).
• Suplementación con buffers u otras sustancias modificadoras del ambiente ruminal (monensina) (Van Nevel y Demeyer, 1995; Nagaraja et al., 1997).
Tabla 3. Efecto del nivel de consumo en las emisiones de metano (Kurihara y Terada, 2001)
A pesar de lo mencionado, los aspectos negativos de la producción de metano en rumen (pérdidas de eficiencia en el uso de la energía por parte de los animales y contribución al efecto invernadero) han motivado un creciente interés en investigar posibles formas de disminuir las emisiones. Desde el punto de vista de la adopción de tecnologías por parte del productor de carne o leche, es claro que un enfoque que se base en las pérdidas de energía dietaria por parte de los animales, que influyen directamente sobre los márgenes económicos que obtiene cada productor, tendrá un efecto mucho más “convincente” que el enfoque de la disminución de la emisión de GEI para evitar el calentamiento global. Czerkawski (1986) concluyó que las pérdidas de eficiencia en el uso de la energía provocadas por las emisiones de metano significaba una pérdida anual de 15.000 millones de libras esterlinas. Cuantificar la contribución de las emisiones de metano por parte de la ganadería al calentamiento global aparece como más dificultoso (Van Nevel y Demeyer, 1995). Sin embargo, un enfoque integrado debería ser tomado en cuenta por gobiernos o formadores de opinión para desarrollar políticas que contribuyan a un incremento en la productividad ganadera en una forma sostenible en el tiempo.
Estrategias para mitigar las emisiones de metano
Está claro que el primer paso para desarrollar estrategias para disminuir las emisiones de metano debería ser lograr estimadores confiables para cuantificar las emisiones por parte de los animales. Al respecto, existen variados métodos, individuales o grupales, que escapan al objetivo de este trabajo pero que pueden ser consultados por el lector interesado (Johnson et al., 1994; Johnson y Johnson, 1995; McCaughey et al., 1997; McGinn et al., 2004). Las guías del Panel Intergubernamental del cambio climático
(IPCC según sus siglas en inglés) contiene una serie de ecuaciones para la cuantificación de las emisiones de metano por parte de la ganadería (IPCC, 2000). En su versión más simplificada, el método IPCC se basa en definir un factor de emisión de metano para una población específica de animales, y multiplicar este factor por el total de animales que componen la población bajo estudio. Una versión un poco más compleja requiere mayor cantidad de datos de requerimientos, consumo y tasa de conversión de metano (expresadas en unidades de energía por cada 100 unidades de energía bruta consumida) de tipos específicos de alimentos.
i) Intervenciones dietarias
Investigaciones de los años ´60 demostraron que la producción de metano
(expresado como porcentaje de la energía del alimento) estaba inversamente relacionada con el nivel de consumo voluntario de alimentos (Blaxter y Clapperton, 1965). Este concepto se ilustra claramente en la Tabla 3, en donde se puede ver que animales consumiendo dietas basadas en silaje de maíz o ryegrass anual, a niveles de mantenimiento o a 3 veces el nivel de mantenimiento, disminuían sus emisiones de metano en un 39 y 22% con silaje de maíz y ryegrass anual, respectivamente (Kurihara y Terada, 2001). Johnson y Johnson (1995) concluyeron que el porcentaje de energía bruta del alimento perdido como metano disminuía 1.6 unidades porcentuales por cada unidad de incremento del consumo por encima del nivel de mantenimiento. Asimismo, la tasa de fermentación de los sustratos es importante: aquellos alimentos que se degradan a tasas más lentas (caso de los carbohidratos estructurales como la celulosa), producen más metano que aquellos que se fermentan a tasas más rápidas (McCrabb, 2002; Eun et al., 2004).
En general, la inclusión de concentrados en dietas de rumiantes trae aparejado una disminución en el pH ruminal, lo que redunda en una disminución de la relación acético:propiónico y de la producción de metano (Van Kessel y Russell, 1996; Lana et al., 1998; Russell, 1998). En el rumen, las bacterias usan metanogénesis o producción de propionato como estrategias para desligarse de poder reductor, siendo la relación entre ambos (metano y propionato) inversa (Lana et al., 1998).
En función de lo expuesto, Van Nevel y Demeyer (1995) concluyeron que toda aquella intervención dietaria (cantidad/calidad) que produjera una alteración en los perfiles de ácidos grasos volátiles producidos a nivel ruminal (más propionato y menos acetato) impactaría sobre la producción de metano. Al respecto, ejemplos de intervenciones incluyen:
• Incrementar el nivel de carbohidratos de alta fermentabilidad en la dieta, en desmedro de forrajes groseros (McCrabb et al., 1998; Eun et al., 2004).
• Tratamientos físicos de forrajes (molido, peleteado) o de granos (tratamientos con vapor) (Zinn, 1987).
• Tratamientos con urea y minerales en dietas de baja calidad (Leng, 1993).
• Suplementación con buffers u otras sustancias modificadoras del ambiente ruminal (monensina) (Van Nevel y Demeyer, 1995; Nagaraja et al., 1997).
Tabla 3. Efecto del nivel de consumo en las emisiones de metano (Kurihara y Terada, 2001)
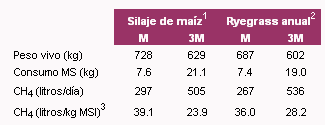
1 Dieta contenía 35% de silaje de maíz, 13% de heno de alfalfa o pastura, y 52% de concentrado.
2 Dieta contenía 35% de ryegrass anual, 15% de heno de alfalfa o pastura, y 50% de concentrado.
3 MSI: Materia seca ingerida.
Investigadores japoneses (Kurihara y Terada, 2001) revisaron bibliografía y encontraron que incrementos en la productividad (ya sea en carne o leche) conllevarían a menores emisiones de metano por unidad de producto (Figuras 2 y 3). Si bien las relaciones son cuadráticas, existe claro potencial en la reducción de emisiones de metano a través del aumento de la productividad.
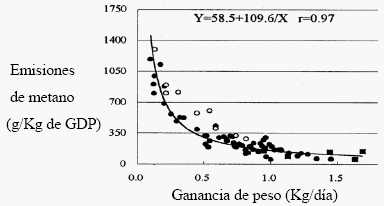
Figura 3. Relación entre la ganancia de peso y la producción de metano para Bos taurus(círculos cerrados) y Bos indicus (círculos abiertos)

Figura 4. Relación entre la producción de leche corregida al 4% de grasa y la producción de metano
ii) Uso de aditivos para reducir las emisiones de metano
Se han investigado variados tipos de aditivos dietarios con el fin de reducir las emisiones de metano. Entre ellos, se pueden citar a los análogos halogenados del metano (Van Nevel y Demeyer, 1995; Dong et al., 1999; Moss et al., 2000), taninos (Puchala et al., 2005); aceptores de hidrógeno que estimulan la producción de ácidos propiónico o butírico (Asanuma et al., 1999; Ungerfeld et al., 2003), extractos de plantas (Lila et al., 2003), galacto-oligosacáridos (Santoso et al., 2004), levaduras (McGinn et al., 2004), etc. También se han desarrollado vacunas específicas contra microorganismos productores de metano, tal lo reportado por McSweeney y McCrabb (2002). Sin embargo, los resultados hasta el momento han sido inconsistentes, y mucha investigación será necesaria antes de poder prescribir con certeza estos tratamientos para reducir las emisiones de metano.
Dentro de los aditivos dietarios que mejores resultados han mostrado, sobresalen los suplementos lipídicos y los antibióticos ionóforos. Los lípidos utilizados como suplementos a las dietas incluyen tanto ácidos grasos de cadena media (Machmüller et al., 2001; Machmüller et al., 2003) o aceites vegetales como el aceite de coco
(Machmüller et al., 1999), girasol (McGinn et al., 2004), pescado (Fievez et al., 2003), u oliva (Ungerfeld et al., 2005). Estos compuestos reducen las emisiones de metano directamente a través de la competencia por equivalentes reductores con los microorganismos metanogénicos y el efecto tóxico directo sobre microorganismos ruminales, e indirectamente a través de la reducción de la población de protozoarios del rumen, que mantienen relaciones simbióticas con los microorganismos metanogénicos. Otros autores (Johnson y Johnson, 1995) atribuyeron esta reducción en las emisiones de metano a una reducción en la cantidad de sustrato fermentable (menor digestibilidad de la fibra). Esta reducción de la digestibilidad de la fibra que resulta del agregado de lípidos por encima de ciertos niveles en la dieta (McGinn et al., 2004) resulta una desventaja para la aplicación de estos aditivos en sistemas de producción basados en aprovechamiento de forrajes.
Potencial de los antibióticos ionóforos para reducir las emisiones de metano
Los antibióticos ionóforos son usados regularmente en países productores de ganado desde hace más de 40 años (Nagaraja, 1995). Dentro de estos antibióticos, el más estudiado es la monensina, desarrollada a partir de investigaciones del Dr. A. P. Raun y colaboradores en los laboratorios Lilly (Raun, et al., 1976; Raun, 1990). Los efectos de la monensina en animales de carne y leche han sido revisados exhaustivamente (Bergen y Bates, 1984; Ipharraguerre y Clark, 2003). Bergen y Bates (1984) concluyeron que la monensina actúa sobre tres aspectos de la fermentación ruminal:
• Un incremento en la producción de propionato y una disminución en la producción de metano, resultando en un incremento en la eficiencia de utilización de la energía consumida por el animal.
• Una reducción en la degradación de la proteína y de la deaminación de los aminoácidos, que resulta en una mejora en el metabolismo proteico.
• Reducción de la producción de ácido láctico y en la formación de espuma en el rumen, lo que reduce la incidencia de desórdenes ruminales tales como acidosis y empaste.
En el presente trabajo nos concentraremos en el potencial de la monensina para reducir las emisiones de metano. La monensina reduce las emisiones de metano debido a que altera las poblaciones microbianas en el rumen, reduciendo las cantidades de bacterias que producen ácidos láctico, acético, butírico, fórmico y gas hidrógeno (susceptibles al ionóforo), favoreciendo a las que producen ácidos propiónico y succínico
(resistentes al ionóforo). Esto trae como resultado una reducción en la relación acético- propiónico (Russell y Strobel, 1989). En un principio, se creyó que la monensina actuaba directamente sobre los microorganismos metanogénicos, pero luego se probó que en realidad la monensina reduce la metanogénesis porque priva a los metanogénicos de hidrógeno y ácido fórmico, los principales precursores del metano en el rumen (McGuffey et al., 2001). Sin embargo, Nagaraja et al. (1997) sostuvieron que la monensina podría inhibir la producción de metano directamente por parte de los microorganismos metanogénicos. El efecto defaunante (reducción de protozoarios) de la monensina puede ser también un factor importante, como ya fuera mencionado anteriormente, aunque esto tendría como contrapartida una posible reducción en la digestibilidad de la fibra (Hino y Asanuma, 2003).
Van Nevel y Demeyer (1995) revisaron una gran cantidad de trabajos y concluyeron que la monensina inhibe la producción de metano in vivo entre un 20 y un
25%, mientras que Schelling (1984) reportó que la reducción puede ser de hasta el 30%. Datos de trabajos in vitro mostraron reducciones de metano de alrededor del 32±2.8% (Van Nevel y Demeyer, 1996). Algunos resultados experimentales in vivo se presentan en la Tabla 4.
Tabla 4. Efecto de la monensina sobre la producción de metano in vivo
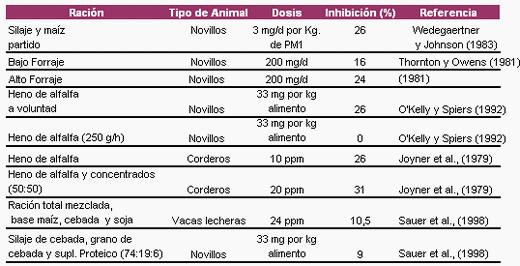
1 PM= Peso metabólico
Sin embargo, algunos autores han sugerido que la reducción en las emisiones de metano por parte de la monensina no persistiría en el tiempo (Rumpler et al., 1986; Johnson et al., 1991; Sauer et al., 1998; Hino y Asanuma, 2003), lo que fue interpretado como una posible adaptación de las poblaciones ruminales al ionóforo (Johnson y Johnson, 1995). Esto es sorprendente porque la alteración de los perfiles de ácidos grasos volátiles (baja relación acético:propiónico) asociada con el agregado de monensina persiste por largos períodos de tiempo (40 a 240 días) (Richardson et al., 1976; Mbanzamihigo et al., 1995; Rogers et., 1997). Estos últimos trabajos no mostraron ninguna evidencia de adaptación microbiana al agregado de la monensina. Desde el punto de vista de la estequiometría de la fermentación ruminal, esto sería posible siempre y cuando no haya otras vías para eliminar el exceso de hidrógeno del rumen (Hino y Asanuma, 2003). Tedeschi et al. (2003) discutieron esta aparente contradicción e indicaron que posiblemente está relacionada con el uso de diferentes métodos para medir la producción de metano, o a las proporciones de forraje y concentrado usadas en diferentes experimentos.
De todas maneras, aún cuando la adaptación de la microflora ruminal existiera, la reducción de las emisiones de metano como resultado del agregado de monensina a las dietas seguiría existiendo, debido a la reducción en el consumo de materia seca que se observa (Nagaraja, 1995). Con respecto a esto, O’Kelly y Spiers (1992) indicaron que la reducción en el consumo voluntario explicaba el 55% de la reducción en las cantidades de metano producidas por novillos.
Conclusiones
La producción de metano es negativa desde un punto de vista productivo y ambiental, y es necesario estudiar formas de decrecer las emisiones. Incrementos en la productividad conllevan a una menor producción de metano. La monensina es un arma para incrementar la productividad, y ha sido probado que reduce las emisiones de metano. El uso de monensina en animales de carne y leche tiene, además de sus efectos positivos sobre la eficiencia productiva, el meteorismo y la cetosis, reduce las emisiones de metano, proporcionando una eficaz arma para reducir el impacto de la ganadería sobre el efecto invernadero. En el futuro, la combinación de monensina con otros productos, tales como los aceptores alternativos de hidrógeno, puede contribuir a disminuir las emisiones de metano en forma más eficiente.
2 Dieta contenía 35% de ryegrass anual, 15% de heno de alfalfa o pastura, y 50% de concentrado.
3 MSI: Materia seca ingerida.
Investigadores japoneses (Kurihara y Terada, 2001) revisaron bibliografía y encontraron que incrementos en la productividad (ya sea en carne o leche) conllevarían a menores emisiones de metano por unidad de producto (Figuras 2 y 3). Si bien las relaciones son cuadráticas, existe claro potencial en la reducción de emisiones de metano a través del aumento de la productividad.
Figura 3. Relación entre la ganancia de peso y la producción de metano para Bos taurus(círculos cerrados) y Bos indicus (círculos abiertos)
Figura 4. Relación entre la producción de leche corregida al 4% de grasa y la producción de metano
ii) Uso de aditivos para reducir las emisiones de metano
Se han investigado variados tipos de aditivos dietarios con el fin de reducir las emisiones de metano. Entre ellos, se pueden citar a los análogos halogenados del metano (Van Nevel y Demeyer, 1995; Dong et al., 1999; Moss et al., 2000), taninos (Puchala et al., 2005); aceptores de hidrógeno que estimulan la producción de ácidos propiónico o butírico (Asanuma et al., 1999; Ungerfeld et al., 2003), extractos de plantas (Lila et al., 2003), galacto-oligosacáridos (Santoso et al., 2004), levaduras (McGinn et al., 2004), etc. También se han desarrollado vacunas específicas contra microorganismos productores de metano, tal lo reportado por McSweeney y McCrabb (2002). Sin embargo, los resultados hasta el momento han sido inconsistentes, y mucha investigación será necesaria antes de poder prescribir con certeza estos tratamientos para reducir las emisiones de metano.
Dentro de los aditivos dietarios que mejores resultados han mostrado, sobresalen los suplementos lipídicos y los antibióticos ionóforos. Los lípidos utilizados como suplementos a las dietas incluyen tanto ácidos grasos de cadena media (Machmüller et al., 2001; Machmüller et al., 2003) o aceites vegetales como el aceite de coco
(Machmüller et al., 1999), girasol (McGinn et al., 2004), pescado (Fievez et al., 2003), u oliva (Ungerfeld et al., 2005). Estos compuestos reducen las emisiones de metano directamente a través de la competencia por equivalentes reductores con los microorganismos metanogénicos y el efecto tóxico directo sobre microorganismos ruminales, e indirectamente a través de la reducción de la población de protozoarios del rumen, que mantienen relaciones simbióticas con los microorganismos metanogénicos. Otros autores (Johnson y Johnson, 1995) atribuyeron esta reducción en las emisiones de metano a una reducción en la cantidad de sustrato fermentable (menor digestibilidad de la fibra). Esta reducción de la digestibilidad de la fibra que resulta del agregado de lípidos por encima de ciertos niveles en la dieta (McGinn et al., 2004) resulta una desventaja para la aplicación de estos aditivos en sistemas de producción basados en aprovechamiento de forrajes.
Potencial de los antibióticos ionóforos para reducir las emisiones de metano
Los antibióticos ionóforos son usados regularmente en países productores de ganado desde hace más de 40 años (Nagaraja, 1995). Dentro de estos antibióticos, el más estudiado es la monensina, desarrollada a partir de investigaciones del Dr. A. P. Raun y colaboradores en los laboratorios Lilly (Raun, et al., 1976; Raun, 1990). Los efectos de la monensina en animales de carne y leche han sido revisados exhaustivamente (Bergen y Bates, 1984; Ipharraguerre y Clark, 2003). Bergen y Bates (1984) concluyeron que la monensina actúa sobre tres aspectos de la fermentación ruminal:
• Un incremento en la producción de propionato y una disminución en la producción de metano, resultando en un incremento en la eficiencia de utilización de la energía consumida por el animal.
• Una reducción en la degradación de la proteína y de la deaminación de los aminoácidos, que resulta en una mejora en el metabolismo proteico.
• Reducción de la producción de ácido láctico y en la formación de espuma en el rumen, lo que reduce la incidencia de desórdenes ruminales tales como acidosis y empaste.
En el presente trabajo nos concentraremos en el potencial de la monensina para reducir las emisiones de metano. La monensina reduce las emisiones de metano debido a que altera las poblaciones microbianas en el rumen, reduciendo las cantidades de bacterias que producen ácidos láctico, acético, butírico, fórmico y gas hidrógeno (susceptibles al ionóforo), favoreciendo a las que producen ácidos propiónico y succínico
(resistentes al ionóforo). Esto trae como resultado una reducción en la relación acético- propiónico (Russell y Strobel, 1989). En un principio, se creyó que la monensina actuaba directamente sobre los microorganismos metanogénicos, pero luego se probó que en realidad la monensina reduce la metanogénesis porque priva a los metanogénicos de hidrógeno y ácido fórmico, los principales precursores del metano en el rumen (McGuffey et al., 2001). Sin embargo, Nagaraja et al. (1997) sostuvieron que la monensina podría inhibir la producción de metano directamente por parte de los microorganismos metanogénicos. El efecto defaunante (reducción de protozoarios) de la monensina puede ser también un factor importante, como ya fuera mencionado anteriormente, aunque esto tendría como contrapartida una posible reducción en la digestibilidad de la fibra (Hino y Asanuma, 2003).
Van Nevel y Demeyer (1995) revisaron una gran cantidad de trabajos y concluyeron que la monensina inhibe la producción de metano in vivo entre un 20 y un
25%, mientras que Schelling (1984) reportó que la reducción puede ser de hasta el 30%. Datos de trabajos in vitro mostraron reducciones de metano de alrededor del 32±2.8% (Van Nevel y Demeyer, 1996). Algunos resultados experimentales in vivo se presentan en la Tabla 4.
Tabla 4. Efecto de la monensina sobre la producción de metano in vivo
1 PM= Peso metabólico
Sin embargo, algunos autores han sugerido que la reducción en las emisiones de metano por parte de la monensina no persistiría en el tiempo (Rumpler et al., 1986; Johnson et al., 1991; Sauer et al., 1998; Hino y Asanuma, 2003), lo que fue interpretado como una posible adaptación de las poblaciones ruminales al ionóforo (Johnson y Johnson, 1995). Esto es sorprendente porque la alteración de los perfiles de ácidos grasos volátiles (baja relación acético:propiónico) asociada con el agregado de monensina persiste por largos períodos de tiempo (40 a 240 días) (Richardson et al., 1976; Mbanzamihigo et al., 1995; Rogers et., 1997). Estos últimos trabajos no mostraron ninguna evidencia de adaptación microbiana al agregado de la monensina. Desde el punto de vista de la estequiometría de la fermentación ruminal, esto sería posible siempre y cuando no haya otras vías para eliminar el exceso de hidrógeno del rumen (Hino y Asanuma, 2003). Tedeschi et al. (2003) discutieron esta aparente contradicción e indicaron que posiblemente está relacionada con el uso de diferentes métodos para medir la producción de metano, o a las proporciones de forraje y concentrado usadas en diferentes experimentos.
De todas maneras, aún cuando la adaptación de la microflora ruminal existiera, la reducción de las emisiones de metano como resultado del agregado de monensina a las dietas seguiría existiendo, debido a la reducción en el consumo de materia seca que se observa (Nagaraja, 1995). Con respecto a esto, O’Kelly y Spiers (1992) indicaron que la reducción en el consumo voluntario explicaba el 55% de la reducción en las cantidades de metano producidas por novillos.
Conclusiones
La producción de metano es negativa desde un punto de vista productivo y ambiental, y es necesario estudiar formas de decrecer las emisiones. Incrementos en la productividad conllevan a una menor producción de metano. La monensina es un arma para incrementar la productividad, y ha sido probado que reduce las emisiones de metano. El uso de monensina en animales de carne y leche tiene, además de sus efectos positivos sobre la eficiencia productiva, el meteorismo y la cetosis, reduce las emisiones de metano, proporcionando una eficaz arma para reducir el impacto de la ganadería sobre el efecto invernadero. En el futuro, la combinación de monensina con otros productos, tales como los aceptores alternativos de hidrógeno, puede contribuir a disminuir las emisiones de metano en forma más eficiente.
Referencias
Asanuma, N., M. Iwamoto, and T. Hino. 1999. Effects of the addition of fumarate on methane production by ruminal microorganisms in vitro. J. Dairy Sci. 82:780-787.
Beauchemin, K. A., and S. M. McGinn. 2005. Methane emissions from feedlot cattle fed barley or corn diets. J. Anim. Sci. 83: 653-661.
Bergen, W. G., and D. B. Bates. 1984. Ionophores: their effect on production efficiency and mode of action. J. Anim. Sci. 58: 1465-1483.
Blaxter, K. L. and J. L. Clapperton. 1965. Prediction of the amount of methane produced by ruminants. Br. J. Nutr. 19: 511-522.
Crutzen, P. J., I. Aselmann, and W. Seiler. 1986. Methane production by domestic animals, wild ruminants, other herbivorous fauna, and humans. Tellus 38B: 271-284.
Czerkawski, J. W. 1986. An introduction to Rumen Studies. Pergamon Press, New York, USA.
Dong, Y., H. D. Bae, T. A. Mc Allister, G. W. Mathison, and K.-J. Cheng. 1999. Effects of exogenous fibrolytic enzymes, á-bromoethanesulfonate and monensin on fermentation in a rumen simulation (RUSITEC) system. Can. J. Anim. Sci. 79:491-498.
Eun, J. S., V. Fellner, and M. L. Gumpertz. 2004. Methane production by mixed ruminal cultures incubated in dual-flow fermentors. J. Dairy Sci. 87: 112-121.
Fievez, V., F. Dohme, M. Danneels, K. Raes, and D. Demeyer. 2003. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Anim. Feed Sci. Technol. 104: 41-58.
Hegarty, R. S. 2002. Strategies for mitigating methane emissions from livestock – Australian options and opportunities. Pages 61-65 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Hino, T., and N. Asanuma. 2003. Suppression of ruminal methanogenesis by decreasing the substrates available to methanogenic bacteria. Nutr. Abstr. Rev. Ser. B Liv. Feeds Feeding 73: 1R-8R.
IPCC (Intergovernmental Panel on Climate Change). 1990. Climate Change: The IPCC science assessment. Cambridge University Press, UK.
IPCC. 2000. Good practice guidance and uncertainly management in national greenhouse gas inventories. Available at http://ipcc-nggip.iges.or.jp/public/gp/gpgaum.htm. Accessed 20/03/2005.
Ipharraguerre, I. R., and J. H. Clark. 2003. Usefulness of ionophores for lactating dairy cows: a review. Anim. Feed Sci. Technol. 106: 39-57.
Johnson, D. E., T. M. Hill, B. R. Carmean, D. W. Lodman, and G. M. Ward. 1991. New perspectives on ruminant methane emissions. Pages 376-379 In Energy Metabolism of Farm Animals. C. Wenk, and M. Boessinger (Eds.). ETH, Zurich, Switzlerland.
Johnson, K. A., M. T. Huyler, H. H. Westberg, B. K. Lamb, and P. Zimmerman. 1994. Measurement of methane emissions from ruminant livestock using a SF6 tracer technique. Environ. Sci. Technol. 28: 359-362.
Johnson, K. A. and D. E. Johnson. 1995. Methane emissions in cattle. J. Anim. Sci. 73: 2483-2492.
Joyner, A. E., L. J. Brown, T. J. Fogg, and R. T. Rossi. 1979. Effect of monensin on growth, feed efficiency and energy metabolism in lambs. J. Anim. Sci. 48: 1065-1069.
Kurihara, M. and F. Terada. 2001. Livestock Production and Greenhouse Gas Emission. Pages 55-68 in Calidad de las Carnes Bovinas Argentinas. Editorial Facultad de Agronomía, Universidad de Buenos Aires.
Lana, R. P., J. B. Russell, and M. E. van Amburgh. 1998. The role of pH in regulating ruminal methane and ammonia production. J. Anim. Sci. 76: 2190-2196.
Leng, R. A. 1993. Quantitative ruminant nutrition- A green science. Aust. J. Agric. Res. 44: 363-380.
Lila, Z. A., N. Mohammed, S. Kanda, T. Komada, and H. Itabashi. 2003. Effect of sarsaponin on ruminal fermentation with particular reference to methane production in vitro. J. Dairy Sci. 86: 3330-3336.
Machmüller, A., and M. Kreuzer. 1999. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 79: 65-72.
Machmüller, A., F. Dohme, C. R. Soliva, M. Wanner, and M. Kreuzer. 2001. Diet composition affects the level of ruminal methane suppression by medium-chain fatty acids. Aust. J. Agric. Res. 52: 713-722.
Machmüller, A., C. R. Soliva, and M. Kreuzer. 2003. Methane-suppressing effect of myristic acid in sheep as affected by dietary calcium and forage production. Brit. J. Nutr. 90: 529-540.
Mbanzamihigo, L., C. J. Van Nevel, and D. Demeyer. 1995. Essai sur l’adaptation fe la fermentation ruminale au monensin. Reprod. Nutr. Dev. 35: 353-365.
McAllister, T. A., E. K. Okine, G. W. Mathison, and K.-J. Cheng. 1996. Dietary, environmental and microbiological aspects of methane production in ruminants. Can. J. Anim. Sci. 76: 231-243.
McCaughey, W. P., K. Wittenberg, and D. Corrigan. 1997. Methane production by steers on pasture. Can. J. Anim. Sci. 77: 519-524.
McCrabb, G. J. 2002. Nutritional options for abatement of methane emissions from beef and dairy systems in Australia. Pages 115-124 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
McCrabb, G. J., M. Kurihara, and R. A. Hunter. 1998. The effect of finishing strategy on lifetime methane production of beef cattle in northern Australia. Proc. of the Aust. Nutr. Soc. 22: 25.
McGinn, S. M, K. A. Beauchemin, T. Coates, and D. Colombatto. 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 82: 3346-3356.
McGuffey, R. K., L. F. Richardson, and J. I. D. Wilkinson. 2001. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci. 84(E-Suppl.): E194-E203.
McSweeney, C. S., and G. J. McCrabb. 2002. Inhibition of rumen methanogenesis and its effects on feed intake, digestion, and animal production. Pages 129-137 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Moss, A. R., J. P. Jouany, and C. J. Newbold. 2000. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 43: 231-253.
Moss, A. R. 2002. Environmental control of methane production by ruminants. Pages 67-76 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Nagaraja, T. G. 1995. Ionophores and antibiotics in ruminants. Pages 173-204 in Biotechnology in Animal Feeds and Animal Feeding. R. J. Wallace, and A. Chesson (Eds.). VCH, Germany.
Nagaraja, T. G., C. J. Newbold, C. J. Van Nevel, and D. I. Demeyer. 1997. Pages 523-632 In The Rumen Microbial Ecosystem. P. N. Hobson, and C. J. Stewart (Eds.). Blackie Academic and Professional, London, UK.
O’Hara, P., J. Freney, and M. Ulyatt. 2003. Abatement of Agricultural Non-Dioxide greenhouse gas emissions. A study of research requirements. Ministry of Agriculture and Forestry. New Zealand.
O’Kelly, J. C., and W. G. Spiers. 1992. Effect of monensin on methane and heat productions of steers fed Lucerne hay. Aust. J. Agric. Res. 43: 1789-1793.
Puchala, R., B. R. Min, A. L. Goetsch, and T. Sahlu. 2005. The effect of a condensed tannin-containing forage on methane emission by goats. J. Anim. Sci. 83: 182-186.
Raun, A. P. 1990. Ionohores – a case study in additive development. Feed Manag. 41: 63.
Raun, A. P., C. O. Cooley, E. L. Potter, R. P. Rathmacher, and L. F. Richardson. 1976. Effect of monensin on feed efficiency of feedlot cattle. J. Anim. Sci. 43: 670-677.
Richardson, L. F., A. P. Raun, E. L. Potter, C. O. Cooley, and R. P. Rathmacher. 1976. Effect of monensin on rumen fermentation in vitro and in vivo. J. Anim. Sci. 43:657-664.
Rogers, M., J. P. Jouany, P. Thivend, and J. P. Fontenot. 1997. The effect of short term and long term monensin supplementation and its subsequent withdrawal on digestion in sheep. Anim. Feed Sci. Technol. 65: 113-127.
Rumpler, W. V., D. E. Johnson, and D. B. Bates. 1986. The effect of high dietary cation concentration on methanogenesis by steers fed diets with and without ionophores. J. Anim. Sci. 62: 1737-1741.
Russell, J. B, and H. J. Strobel. 1989. The effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 55: 1-6.
Russell, J. B. 1998. The importance of pH in the regulation of ruminal acetate and propionate ratio and methane production in vitro. J. Dairy Sci. 81: 3222-3230.
Santoso, B., B. Mwenya, C. Sar, Y. Gamo, T. Kobayashi, R. Morikawa, K. Kimura, H. Mizukoshi, and J. Takahashi. 2004. Effects of supplementing galacto- oligosaccharides, Yucca schidigera or nisin on rumen methanogenesis, nitrogen and energy metabolism in sheep. Liv. Prod. Sci. 91: 209-217.
Sauer, F. D., V. Fellner, R. Kinsman, J. K. G. Kramer, H. A. Jackson, A. J. Lee, and S. Chen. 1998. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added to the diet. J. Anim. Sci. 76: 906-914.
Schelling, G. T. 1984. Monensin mode of action in the rumen. J. Anim. Sci. 58: 1518-1527.
Tedeschi, L. O., D. G. Fox, and T. P. Tylutki. 2003. Potential environmental benefits of ionophores in ruminant diets. J. Environ. Qual. 32: 1591-1602.
Thornton, J. H., and F. N. Owens. 1981. Monensin supplementation and in vivo methane production by steers. J. Anim. Sci. 52: 628-634.
Ungerfeld, E. M., S. R. Rust, and R. J. Burnett. 2003. Effects of combinations of propyonic acid and crotonic acid or 3-butenoic acid on ruminal digestibility and microbial efficiency in vitro. Page 37 in Proc. Gastr. Function Conf. Chicago, IL, USA.
Ungerfeld, E. M., S. R. Rust, R. J. Burnett, M. T. Yokoyama, and J. K. Wang. 2005. Effects of two lipids on in vitro ruminal methane production. Anim. Feed Sci. Technol. 119: 179-185.
US EPA (United States Environmental Policy Agency). 1994. International Anthropogenic methane emissions: Estimates for 1990. US EPA 230-R-93-010. Washington, DC, USA.
Van Kessel, J. A. S. and J. B. Russell. 1996. The effect of pH on ruminal methanogenesis. FEMS Microbiol. Ecol. 20: 205-210.
Van Nevel, C., and D. Demeyer. 1995. Feed additives and other interventions for decreasing methane emissions. Pages 329-349 in Biotechnology in Animal Feeds and Animal Feeding. R. J. Wallace, and A. Chesson (Eds.). VCH, Germany.
Van Nevel, C., and D. Demeyer. 1996. Control of rumen methanogenesis. Environ. Monit. Assess. 42: 73-97.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. Cornell University Press, Ithaca, NY, USA.
Wedegaertner, T. C., and D. E. Johnson. 1983. Monensin effects on digestibility, methanogenesis and heat increment of a cracked corn- silage fed to steers. J. Anim. Sci. 57: 168-177.
Wolin, M. J., and T. L. Miller. 1988. Microbe interactions in the rumen microbial ecosystem. Pages 343-359 In The Rumen Microbial Ecosystem. P. N. Hobson, (Ed.). Elsevier Applied Science, New York, USA.
Zinn, R. A. 1987. Influence of lasalocid and monensin plus tylosin on comparative feeding value of steam-flaked versus dry-rolled corn in diets for feedlot cattle. J. Anim. Sci. 65: 256-266.
Asanuma, N., M. Iwamoto, and T. Hino. 1999. Effects of the addition of fumarate on methane production by ruminal microorganisms in vitro. J. Dairy Sci. 82:780-787.
Beauchemin, K. A., and S. M. McGinn. 2005. Methane emissions from feedlot cattle fed barley or corn diets. J. Anim. Sci. 83: 653-661.
Bergen, W. G., and D. B. Bates. 1984. Ionophores: their effect on production efficiency and mode of action. J. Anim. Sci. 58: 1465-1483.
Blaxter, K. L. and J. L. Clapperton. 1965. Prediction of the amount of methane produced by ruminants. Br. J. Nutr. 19: 511-522.
Crutzen, P. J., I. Aselmann, and W. Seiler. 1986. Methane production by domestic animals, wild ruminants, other herbivorous fauna, and humans. Tellus 38B: 271-284.
Czerkawski, J. W. 1986. An introduction to Rumen Studies. Pergamon Press, New York, USA.
Dong, Y., H. D. Bae, T. A. Mc Allister, G. W. Mathison, and K.-J. Cheng. 1999. Effects of exogenous fibrolytic enzymes, á-bromoethanesulfonate and monensin on fermentation in a rumen simulation (RUSITEC) system. Can. J. Anim. Sci. 79:491-498.
Eun, J. S., V. Fellner, and M. L. Gumpertz. 2004. Methane production by mixed ruminal cultures incubated in dual-flow fermentors. J. Dairy Sci. 87: 112-121.
Fievez, V., F. Dohme, M. Danneels, K. Raes, and D. Demeyer. 2003. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Anim. Feed Sci. Technol. 104: 41-58.
Hegarty, R. S. 2002. Strategies for mitigating methane emissions from livestock – Australian options and opportunities. Pages 61-65 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Hino, T., and N. Asanuma. 2003. Suppression of ruminal methanogenesis by decreasing the substrates available to methanogenic bacteria. Nutr. Abstr. Rev. Ser. B Liv. Feeds Feeding 73: 1R-8R.
IPCC (Intergovernmental Panel on Climate Change). 1990. Climate Change: The IPCC science assessment. Cambridge University Press, UK.
IPCC. 2000. Good practice guidance and uncertainly management in national greenhouse gas inventories. Available at http://ipcc-nggip.iges.or.jp/public/gp/gpgaum.htm. Accessed 20/03/2005.
Ipharraguerre, I. R., and J. H. Clark. 2003. Usefulness of ionophores for lactating dairy cows: a review. Anim. Feed Sci. Technol. 106: 39-57.
Johnson, D. E., T. M. Hill, B. R. Carmean, D. W. Lodman, and G. M. Ward. 1991. New perspectives on ruminant methane emissions. Pages 376-379 In Energy Metabolism of Farm Animals. C. Wenk, and M. Boessinger (Eds.). ETH, Zurich, Switzlerland.
Johnson, K. A., M. T. Huyler, H. H. Westberg, B. K. Lamb, and P. Zimmerman. 1994. Measurement of methane emissions from ruminant livestock using a SF6 tracer technique. Environ. Sci. Technol. 28: 359-362.
Johnson, K. A. and D. E. Johnson. 1995. Methane emissions in cattle. J. Anim. Sci. 73: 2483-2492.
Joyner, A. E., L. J. Brown, T. J. Fogg, and R. T. Rossi. 1979. Effect of monensin on growth, feed efficiency and energy metabolism in lambs. J. Anim. Sci. 48: 1065-1069.
Kurihara, M. and F. Terada. 2001. Livestock Production and Greenhouse Gas Emission. Pages 55-68 in Calidad de las Carnes Bovinas Argentinas. Editorial Facultad de Agronomía, Universidad de Buenos Aires.
Lana, R. P., J. B. Russell, and M. E. van Amburgh. 1998. The role of pH in regulating ruminal methane and ammonia production. J. Anim. Sci. 76: 2190-2196.
Leng, R. A. 1993. Quantitative ruminant nutrition- A green science. Aust. J. Agric. Res. 44: 363-380.
Lila, Z. A., N. Mohammed, S. Kanda, T. Komada, and H. Itabashi. 2003. Effect of sarsaponin on ruminal fermentation with particular reference to methane production in vitro. J. Dairy Sci. 86: 3330-3336.
Machmüller, A., and M. Kreuzer. 1999. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 79: 65-72.
Machmüller, A., F. Dohme, C. R. Soliva, M. Wanner, and M. Kreuzer. 2001. Diet composition affects the level of ruminal methane suppression by medium-chain fatty acids. Aust. J. Agric. Res. 52: 713-722.
Machmüller, A., C. R. Soliva, and M. Kreuzer. 2003. Methane-suppressing effect of myristic acid in sheep as affected by dietary calcium and forage production. Brit. J. Nutr. 90: 529-540.
Mbanzamihigo, L., C. J. Van Nevel, and D. Demeyer. 1995. Essai sur l’adaptation fe la fermentation ruminale au monensin. Reprod. Nutr. Dev. 35: 353-365.
McAllister, T. A., E. K. Okine, G. W. Mathison, and K.-J. Cheng. 1996. Dietary, environmental and microbiological aspects of methane production in ruminants. Can. J. Anim. Sci. 76: 231-243.
McCaughey, W. P., K. Wittenberg, and D. Corrigan. 1997. Methane production by steers on pasture. Can. J. Anim. Sci. 77: 519-524.
McCrabb, G. J. 2002. Nutritional options for abatement of methane emissions from beef and dairy systems in Australia. Pages 115-124 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
McCrabb, G. J., M. Kurihara, and R. A. Hunter. 1998. The effect of finishing strategy on lifetime methane production of beef cattle in northern Australia. Proc. of the Aust. Nutr. Soc. 22: 25.
McGinn, S. M, K. A. Beauchemin, T. Coates, and D. Colombatto. 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 82: 3346-3356.
McGuffey, R. K., L. F. Richardson, and J. I. D. Wilkinson. 2001. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci. 84(E-Suppl.): E194-E203.
McSweeney, C. S., and G. J. McCrabb. 2002. Inhibition of rumen methanogenesis and its effects on feed intake, digestion, and animal production. Pages 129-137 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Moss, A. R., J. P. Jouany, and C. J. Newbold. 2000. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 43: 231-253.
Moss, A. R. 2002. Environmental control of methane production by ruminants. Pages 67-76 in Greenhouse Gases and Animal Agriculture. Takahashi, J., and B. A. Young, (Eds.). Elsevier, Amsterdam, The Netherlands.
Nagaraja, T. G. 1995. Ionophores and antibiotics in ruminants. Pages 173-204 in Biotechnology in Animal Feeds and Animal Feeding. R. J. Wallace, and A. Chesson (Eds.). VCH, Germany.
Nagaraja, T. G., C. J. Newbold, C. J. Van Nevel, and D. I. Demeyer. 1997. Pages 523-632 In The Rumen Microbial Ecosystem. P. N. Hobson, and C. J. Stewart (Eds.). Blackie Academic and Professional, London, UK.
O’Hara, P., J. Freney, and M. Ulyatt. 2003. Abatement of Agricultural Non-Dioxide greenhouse gas emissions. A study of research requirements. Ministry of Agriculture and Forestry. New Zealand.
O’Kelly, J. C., and W. G. Spiers. 1992. Effect of monensin on methane and heat productions of steers fed Lucerne hay. Aust. J. Agric. Res. 43: 1789-1793.
Puchala, R., B. R. Min, A. L. Goetsch, and T. Sahlu. 2005. The effect of a condensed tannin-containing forage on methane emission by goats. J. Anim. Sci. 83: 182-186.
Raun, A. P. 1990. Ionohores – a case study in additive development. Feed Manag. 41: 63.
Raun, A. P., C. O. Cooley, E. L. Potter, R. P. Rathmacher, and L. F. Richardson. 1976. Effect of monensin on feed efficiency of feedlot cattle. J. Anim. Sci. 43: 670-677.
Richardson, L. F., A. P. Raun, E. L. Potter, C. O. Cooley, and R. P. Rathmacher. 1976. Effect of monensin on rumen fermentation in vitro and in vivo. J. Anim. Sci. 43:657-664.
Rogers, M., J. P. Jouany, P. Thivend, and J. P. Fontenot. 1997. The effect of short term and long term monensin supplementation and its subsequent withdrawal on digestion in sheep. Anim. Feed Sci. Technol. 65: 113-127.
Rumpler, W. V., D. E. Johnson, and D. B. Bates. 1986. The effect of high dietary cation concentration on methanogenesis by steers fed diets with and without ionophores. J. Anim. Sci. 62: 1737-1741.
Russell, J. B, and H. J. Strobel. 1989. The effect of ionophores on ruminal fermentation. Appl. Environ. Microbiol. 55: 1-6.
Russell, J. B. 1998. The importance of pH in the regulation of ruminal acetate and propionate ratio and methane production in vitro. J. Dairy Sci. 81: 3222-3230.
Santoso, B., B. Mwenya, C. Sar, Y. Gamo, T. Kobayashi, R. Morikawa, K. Kimura, H. Mizukoshi, and J. Takahashi. 2004. Effects of supplementing galacto- oligosaccharides, Yucca schidigera or nisin on rumen methanogenesis, nitrogen and energy metabolism in sheep. Liv. Prod. Sci. 91: 209-217.
Sauer, F. D., V. Fellner, R. Kinsman, J. K. G. Kramer, H. A. Jackson, A. J. Lee, and S. Chen. 1998. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added to the diet. J. Anim. Sci. 76: 906-914.
Schelling, G. T. 1984. Monensin mode of action in the rumen. J. Anim. Sci. 58: 1518-1527.
Tedeschi, L. O., D. G. Fox, and T. P. Tylutki. 2003. Potential environmental benefits of ionophores in ruminant diets. J. Environ. Qual. 32: 1591-1602.
Thornton, J. H., and F. N. Owens. 1981. Monensin supplementation and in vivo methane production by steers. J. Anim. Sci. 52: 628-634.
Ungerfeld, E. M., S. R. Rust, and R. J. Burnett. 2003. Effects of combinations of propyonic acid and crotonic acid or 3-butenoic acid on ruminal digestibility and microbial efficiency in vitro. Page 37 in Proc. Gastr. Function Conf. Chicago, IL, USA.
Ungerfeld, E. M., S. R. Rust, R. J. Burnett, M. T. Yokoyama, and J. K. Wang. 2005. Effects of two lipids on in vitro ruminal methane production. Anim. Feed Sci. Technol. 119: 179-185.
US EPA (United States Environmental Policy Agency). 1994. International Anthropogenic methane emissions: Estimates for 1990. US EPA 230-R-93-010. Washington, DC, USA.
Van Kessel, J. A. S. and J. B. Russell. 1996. The effect of pH on ruminal methanogenesis. FEMS Microbiol. Ecol. 20: 205-210.
Van Nevel, C., and D. Demeyer. 1995. Feed additives and other interventions for decreasing methane emissions. Pages 329-349 in Biotechnology in Animal Feeds and Animal Feeding. R. J. Wallace, and A. Chesson (Eds.). VCH, Germany.
Van Nevel, C., and D. Demeyer. 1996. Control of rumen methanogenesis. Environ. Monit. Assess. 42: 73-97.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. Cornell University Press, Ithaca, NY, USA.
Wedegaertner, T. C., and D. E. Johnson. 1983. Monensin effects on digestibility, methanogenesis and heat increment of a cracked corn- silage fed to steers. J. Anim. Sci. 57: 168-177.
Wolin, M. J., and T. L. Miller. 1988. Microbe interactions in the rumen microbial ecosystem. Pages 343-359 In The Rumen Microbial Ecosystem. P. N. Hobson, (Ed.). Elsevier Applied Science, New York, USA.
Zinn, R. A. 1987. Influence of lasalocid and monensin plus tylosin on comparative feeding value of steam-flaked versus dry-rolled corn in diets for feedlot cattle. J. Anim. Sci. 65: 256-266.
Temas relacionados
Autores:

Únete para poder comentar.
Una vez que te unas a Engormix, podrás participar en todos los contenidos y foros.
* Dato obligatorio
¿Quieres comentar sobre otro tema? Crea una nueva publicación para dialogar con expertos de la comunidad.
Crear una publicación
CADIA - Centro Argentino de Ingenieros Agrónomos
25 de junio de 2016
Las super bacterias, son producto escencialmente, del pésimo uso que se hace de los antibióticos de uso humano. Se prescriben tratamientos con antibióticos y quimioterápicos, por la aparición de infecciones de poca importancia. Los pacientes abandonan los tratamientos a la mitad. Se utilizan dosis muy bajas, que solo retienen a los patógenos, favoreciendo la aparición de formas resistentes. La tendencia mundial es a no utilizar antibióticos de uso humano en animales. En general los Ionóforos no son usados para humanos, tampoco macrólidos como la tylosina.
En términos generales, hay una creciente satanización del uso de estos recursos terapéuticos, pero paralelamente, se pretenden alimentos de bajo costo. Los bajos costos se obtienen por aplicación de técnicas de confinamiento. Inevitablemente, si confinamos cerdos, pollos, terneros o lo que sea, en pequeños espacios, los problemas sanitarios estarán golpeandonos a la puerta.
Los mecanismos por los cuales, cualquier técnica, producto o práctica de producción destinada a controlar a los organismos indeseables, tendrá como respuesta, la contramedida o respuesta de generación de resistencias por parte del organismo a combatir. NO DIFIERE MAYORMENTE, DE LA APARICIÓN DE MALEZAS RESISTENTES AL GLIFOSATO, LUEGO DE MUCHOS AÑOS DE MONOCULTIVO Y APLICACIÓN DE ESE HERBICIDA SOBRE UN MISMO LOTE.

Universidad Autónoma Chapingo
24 de junio de 2016
FRANCESA de EUA señala un punto clave en el orden de contaminantes. Si el ganado o fauna silvestre no consume esos pastos de las praderas y hojarasca del bosque, los mismos caen al suelo y serán descompuestos por las bacterias y hongos del suelo.
Siguiendo el ciclo del Carbono (CHON) se le puede dar seguimiento a la producción de gas metano. Se vera que es un ciclo NATURAL.
Ahora muchos de los cálculos están basados en POTENCIALES de fermentación de la materia orgánica en cámaras cerradas de evaluación y en REALIDAD mucha de la materia orgánica queda almacenada en MATERIA ORGÁNICA, carne y excrementos o mejoradores de la fertilidad del suelo, NO TODO ES FERMENTADO COMPLETAMENTE en metano, solo una pequeña fracción.
DE NO SER ASÍ Y SI SE SATANIZA LOS ANIMALES EN CONFINAMIENTO YA ESTARÍAN MUERTOS, llámese engordas y establos.
Es decir si los animales no lo consumen de alguna manera pasan por el ciclo biológico y en esos procesos se produce gas metano CH4.


24 de junio de 2016
Estimados foristas:
Todos estamos de acuerdo en los beneficios de los ionoferos en la reducción de metano excretado al medio ambiete en aquellos animals que lo reciben en su dieta diaria.
Sin embargo, tomemos en cuenta que esta es una ventaja indirecta que los productos comerciales basados en este antibiótico ofrecen.
La intención inicial directa de esta tecnología 20 anos atrás,no daba un comino acerca del medio ambiente, sus intenciones eran claramente económicas y basadas en una mayor produccion de leche, lana, carne,etc, con una reduccion en consumo de material seca.
La resurgencia en la preocupación de la contaminación atmosférica con gases altos en carbón, ignorada por los países industriales, aun después del tratado de Kioto, ha abierto una gran oportunidad economica para aquellas compañías que poseen tecnologías y productos para otros fines comerciales; pero. que podrían adaptarse muy bien para este nuevo nicho economico.
Elanco y otros no son la excepción y hoy por hoy su lema comercial en produccion animal, no es el mismo de hace 20 anos, se acuerdan: mas leche con menor consume de material seca". Hoy es la reducción del metano excretado y eructado por los rumiantes que lo reciben en su dieta.
Hagamos un poco de historia, el metano siempre ha existido en la atmósfera de nuestro planeta, en cantidades tales y junto con otros gases nocivos que no era posible la existencia de vida otra que no fuera en el agua (océanos, lagos y rios).
Tomaron millones de anos para que las concentraciones de esos gases en la atmósfera fueran reducidos a los niveles actuales.
Voy a specular aquí un poco y uds me corrigen si estoy equivocado y acerca de poblaciones demograficas de rumiantes antes del descubrimiento de América:
La población de rumiantes en el planeta era mayor o menor que la existente hoy en dia?
Era la contaminación atmosférica mayor en esos dias comparada con la existente hoy dia?
La respuesta sin temor a equivocarme es que antes del descubrimiento de América habían mas rumiantes en el planeta, la mayoría en estado salvaje y por lo tanto en pastoreo natural, eructando enormes cantidades de metano al medio ambiente. En lo que es USA y Canadá se calcula que habían al menos 50 millones de cabezas de bisontes. En el continente Africano los millones de rumiantes que habían es incalculable.
Ahora, sin duda alguna la calidad del aire que se respiraba en esos dias era mucho mejor que la existente en Mexico DF o Beiging.


23 de junio de 2016
Si tal efecto: reducción del metano. Excelente aporte al efecto indeseable del gas invernadero (GI. Como se sabe, ahora los rumiantes tiene un rol importante como fuente de GI (http://mrojas.perulactea.com/2014/12/09/ciencia-veterinaria-y-calentamiento-global-una-mini-vision-peruana-a-proposito-de-la-cop20/#more-1948). Al respecto, será bueno culturizar la utilización del forraje, en el momento preciso de "optimo valor alimenticio". Cuanto mayor edad del forraje, mayor cantidad de lignina, y también mayor fuente de GI.

Universidad de Buenos Aires
22 de junio de 2016
Gracias a todos los que han comentado. Aclaro que no tengo relación comercial con las empresas que producen o venden monensina. Si bien tengo un costado científico/investigativo y docente, además realizo actividad privada en Argentina, Chile, Uruguay, México, Guatemala, Colombia y Brasil, con consultorías en diversos sistemas de producción. Como técnico, mi cliente es el productor o la empresa de pre-mezclas pero siempre diferenciado de la venta de los productos. Recomiendo lo que he probado o investigado.
Saludos cordiales
Universidad de Buenos Aires
22 de junio de 2016
Señores del foro
Veo con interés que hay un renovado número de opiniones sobre algo que escribí en 2005. Tomaría con mucha cautela las expresiones del forista Gómez López, que relaciona el uso de monensina con la aparición de súper bacterias. La aparición de súper bacterias está relacionada con la automedicación excesiva en humanos, de antibióticos usados en MEDICINA HUMANA. No hay evidencia seria científica con respecto al uso de monensina, antibiótico que molecularmente no tiene nada que ver con los antibóticos usados en humanos.
Iré contestando otras opiniones, que por cierto son siempre válidas, pero rogaría que hablemos con datos de ciencia y no con creencias.
Saludos

Universidad Autónoma Chapingo
22 de junio de 2016
Muy interesante el FORO de discusión.
Los Pros y Contras enriquecen los comentarios y cierto que hay tendencias comerciales para favorecer o rechazar productos, pero el pique es bueno para que se de apertura a la opinión de otros foristas.
CADIA - Centro Argentino de Ingenieros Agrónomos
22 de junio de 2016
EXCELENTE CONTRIBUCIÓN.
Lo que no debemos perder de vista siempre, es que muchas restricciones son a menudo "barreras para arancelarias".
Tienen como objetivo, disminuir la competitividad de nuestros productos u obligarnos a aumentar nuestros costos de producción (para lo que los Argentinos no necesitamos ayuda externa).
Les relato una experiencia propia de los años 70`s.
Hicimos las primeras experiencias locales usando cama de pollos parrilleros como el 40 % de las raciones, con el agregado de un poco de expeller de girasol y sorgo.
Comparando con raciones constituidas con afrechillos, de trigo y maiz, minerales, sorgo y expeller de maní, pese a ser, a todas luces, raciones más adecuadas, con mejores datos de composición centesimal, LA RACION CON CASI LA MITAD DE CAMA DE POLLO, SACABA UNA IMPORTANTE VENTAJA. Tardamos en advertirlo, la razón era el contenido de MONENSINA, incluida en los alimentos que comían los pollos como "coccidicida", de la marca "Elanco ban 100". Nuestros funcionarios de SENASA, se rinden a las comisiones negociadoras europeas. Le resulta más sencillo que presionar con alternativas negociadoras de "tu me quitas, yo te quito". Por el momento el uso de ionóforos es la mejor alternativa para disminuir las emisiones de CH4 y mejorar las conversiones alimentarias.

22 de junio de 2016
Esta recopilación que el ing. Colombatto, esta completamente apegada a la mayoría de los estudios y experimentos efectuados en aquellas escuelas de nutrición animal alrededor del mundo. Criticismo al mismo con argumentos que no tienen ningún respaldo científico es puramente especulación y debería preguntarse al que la hace que muestre sus referencias científicas.
Este documento señala claramente los mecanismos de accion de los Ionoferos y sus limitaciones cuando se emplean en animales alimentados con raciones que difieren en composición de nutrientes.
Me parece a mi que esta mas indicado su uso en animales bajo dietas en donde los cereales y ensilajes forman una mayor parte de la ración, con la intención de obtener mas acida propiónico.
Sin embargo, mis dudas acerca de sus beneficios en animales en pastoreo, en donde las dietas de los mismos son mas altas en forrajes,
Paradoxicamente, seria este tipo de explotaciones las que mas se beneficiarían con esta tecnología, ya que son los ruminales en pastoreo, los que contribuyen en mayor cantidad de emisiones al medio ambiente.
Menciono a continuación tres puntos que me llamaron la atención del trabajo citado:
Otros autores (Johnson y Johnson, 1995) atribuyeron esta reducción en las emisiones de metano a una reducción en la cantidad de sustrato fermentable (menor digestibilidad de la fibra). Esta reducción de la digestibilidad de la fibra que resulta del agregado de lípidos por encima de ciertos niveles en la dieta (McGinn et al., 2004) resulta una desventaja para la aplicación de estos aditivos en sistemas de producción basados en aprovechamiento de forrajes.
Reducción de la producción de ácido láctico y en la formación de espuma en el rumen, lo que reduce la incidencia de desórdenes ruminales tales como acidosis y empaste.
De todas maneras, aún cuando la adaptación de la microflora ruminal existiera, la reducción de las emisiones de metano como resultado del agregado de monensina a las dietas seguiría existiendo, debido a la reducción en el consumo de materia seca que se observa (Nagaraja, 1995). Con respecto a esto, O’Kelly y Spiers (1992) indicaron que la reducción en el consumo voluntario explicaba el 55% de la reducción en las cantidades de metano producidas por novillos


31 de mayo de 2016
Sin problema he leído muchos artículos sobre este tema que no es nuevo y si hay granjas donde tenemos años utilizando monensina, solo quise que supieras que también hay artículos donde explican la contraparte que comentó el Ing. Iván.
gracias



Te puede interesar
.jpg&w=3840&q=75)